EL MÉTODO CIENTÍFICO
ES UN PROCEDIMIENTO PARA DESCRIBIR LAS CONDICIONES EN QUE SE PRESENTA SUCESOS, ESPECÍFICOS , VITRIFICABLE DE RAZONAMIENTOS RIGUROSOS Y UNA OBSERVACIÓN EMPÍRICA
ETAPAS PARA REALIZAR UN MÉTODO CIENTÍFICO:
- IDEAS ,OBSERVACIONES
- RECONOCIMIENTO DE PROBLEMAS Y EVALUACIÓN DE EVIDENCIA
- FORMULACIÓN DE HIPÓTESIS: GENERACIÓN DE SOLUCIONES DE CREATIVO Y LÓGICA
- FORMULACIÓN DE OBJETIVOS Y MÉTODOS, EXPERIMENTOS CONTROLADOS
- PRUEBA DE HIPÓTESIS, EXPERIMENTACIÓN , RECOLECCIÓN DE DATOS
- JUICIO Y CONCLUSIONES SOBRE LOS PROCEDIMIENTOS ,RESULTADOS Y TEORÍAS EN COMPARACIÓN DE RESULTADOS CON HIPÓTESIS


BIBLIOGRAFIA:http://myprofeciencias.wordpress.com/2011/01/13/el-mtodo-cientfico/
MATERIA
ES TODO LO QUE TIENE MASA Y VOLUMEN
MASA : ES AL CANTIDAD DE MATERIA QUE POSEE UN CUERPO .SE MIDE EN KILOGRAMOS, GRAMOS, MILIGRAMOS
VOLUMEN : CANTIDAD DE ESPACIO QUE OCUPA LA MATERIA QUE COMPONE UN CUERPO .SE MIDE EN METROS CUBICO, LITRO, CENTÍMETROS CÚBICOS , MILILITROS
AQUÍ SE LE PRESENTARA LAS CLASIFICACIONES DE LA MATERIA




 Esta interacción se produce por las atracciones electrostáticas que se producen entre la zona cargada negativamente de una molécula y la positiva de otra, lo que provoca que las moléculas se vayan orientando unas con respecto a otras.
Esta interacción se produce por las atracciones electrostáticas que se producen entre la zona cargada negativamente de una molécula y la positiva de otra, lo que provoca que las moléculas se vayan orientando unas con respecto a otras.








METABOLISMO
INFINIDADES DE REACCIONES QUÍMICA QUE OCURRE DENTRO DE LAS CÉLULAS PARA EL DESARROLLO DE SU PROCESOS VITALES
FUNCIONES
- DE DIGESTIÓN : SE DA LA ENERGÍA
- CONTRIBUCIÓN : DE BIOMOLECULAS
- ELIMINACION , REGULACIÓN, EQUILIBRIO: DE FORMACIÓN , RENOVACIÓN, DE ESTRUCTURAS
- REPORDUCCION: ES EL MANTENIMIENTO DE LA ESPECIE
CLASIFICACIÓN
- ANABOLISMO : SÍNTESIS O FORMACIÓN DE SUSTANCIAS
- SE DAN DE PEQUEÑAS O SIMPLES HASTA GRANDES O COMPLEJAS
- GENERAN ENERGÍA ENDERGONICA NO EXPONTANIA
- EJEMPLO LAS PROTEÍNAS
aa-aa-aa-aa-aa= proteinas
2. CATABOLISMO : DESTRUCCIÓN DE SUSTANCIAS
- DA DE GRANDES O COMPLEJAS HASTA PEQUEÑAS O SIMPLES
- LIBERAN ENERGIA DE EXERGONICA SON EXPOTANIAS
- EJEMPLO : LA LACTOS = GALACTODA B + GLUCOSA + ENERGIA
ANTILOLICO : DA DE ANABOLICO HASTA CATABOLICO
ANAPLEUROTICO: DA DE CATABOLICO HASTA ANABOLICO
- AEROBIO: PRECIENCIA DE CO2 Y ENERGIA
- ANAEREBICO: ACIDO LACTOCO ,UNSENCIA DE O2
TRANSPORTE DE ENERGÍA ATP
- ES UN NUCLEOTICO ENERGÉTICO
- AZÚCAR ES LA RIBOSOMA ES DE FORMA DE ANILLO
- BASES NITROGENADAS LA ADENINA SON 2 ANILLOS
- FOSFATOS SON 3 GRUPOS
- C10,H16,N5,O13,P3
CLASIFICADOS
- NAD = 3 ATP
- FAD = 2 ATP
PROCESOS DONDE SE ENCUENTRE ATP
- ANABOLISMO: BIOSINTESIS, LIPIDOS
- TRANSPORTE ACTIVO : DE LA MEMBRANA PLASMÁTICA DE LA ENERGÍA A PROTEÍNA INTEGRAL TRANSPORTADORAS
- CONTRACCIÓN DE MÚSCULOS
- CICLO CELULAR QUE SON MEIOSIS Y MITOSIS
Enzimas
Las
enzimas son proteínas que catalizan todas las reacciones
bioquímicas. Además de su importancia como catalizadores biológicos, tienen
muchos usos médicos y comerciales.
Un catalizador es una
sustancia que disminuye la energía de activación de una reacción química. Al
disminuir la energía de activación, se incrementa la velocidad de la reacción.
La
mayoría de las reacciones de los sistemas vivos son reversibles, es decir, que
en ellas se establece el equilibrio químico. Por lo tanto, las enzimas aceleran
la formación de equilibrio químico, pero no afectan las concentraciones finales
del equilibrio.
De
acuerdo a su complejidad las enzimas se clasifican como:
En
las proteínas conjugadas podemos distinguir dos partes:
- Apoenzima: Es la parte polipeptídica de la enzima.
- Cofactor: Es la parte no proteica de la enzima.
La
combinación de la apoenzima y el cofactor forman la holoenzima.
Los
cofactores pueden ser:
*Iones metálicos: Favorecen la
actividad catalítica general de la enzima, si no están presentes, la enzima no
actúa. Estos iones metálicos se denominanactivadores. Ejemplos: Fe2+,
Mg2+, Cu2+, K+, Na+ y Zn2+
*La
mayoría de los otros cofactores son coenzimas las cuales
generalmente son compuestos orgánicos de bajo peso
molecular, por ejemplo, las vitaminas del complejo “B” son coenzimas que se requieren para
una respiración celular adecuada.
Tipo de enzimas
|
Actividad
|
Hidrolasas
|
Catalizan reacciones de hidrólisis. Rompen las
biomoléculas con moléculas de agua. A este tipo pertenecen las enzimas
digestivas.
|
Isomerasas
|
Catalizan las reacciones en las cuales un isómero se
transforma en otro, es decir, reacciones de
isomerización.
|
Ligasas
|
Catalizan la unión de moléculas.
|
Liasas
|
Catalizan las reacciones de adición de enlaces o
eliminación, para producir dobles enlaces.
|
Oxidorreductasas
|
Catalizan reacciones de óxido-reducción.
Facilitan latransferencia de electrones de una molécula a
otra.Ejemplo; la
glucosa, oxidasa cataliza la oxidación de glucosa a ácido glucónico.
|
Tansferasas
|
Catalizan la transferencia de un
grupo de una sustancia a otra. Ejemplo: la transmetilasa es
una enzima que cataliza la transferencia de un grupo metilo de una molécula a
otra.
|
La sustancia sobre la cual actúa una
enzima se llama sustrato.
Los
sustratos son específicos para cada enzima:
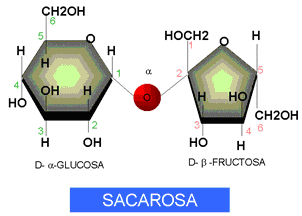
La sacarosa es el sustrato de la sacarasa que actúa rompiéndola en sus componentes.
La sacarosa es el sustrato de la sacarasa que actúa rompiéndola en sus componentes.
Las
enzimas actúan de acuerdo con la siguiente secuencia: La enzima (E) y
el sustrato
(S) se combinan para formar un complejo intermedio enzima sustrato (E-S), el cual se descompone formando un producto y regenerando la enzima.
(S) se combinan para formar un complejo intermedio enzima sustrato (E-S), el cual se descompone formando un producto y regenerando la enzima.
El
grado de especificidad de las enzimas es muy alto, pueden distinguir incluso
entre diferentes tipos de isómeros. Se cree que la especificidad de la enzima
es debido a la forma particular de una pequeña parte conocida como sitio activo, la cual se fija a
la contraparte complementaria en el sustrato.
Concentración del sustrato.- A mayor concentración del
sustrato, a una concentración fija de la enzima se obtiene la velocidad máxima.
Después de que se alcanza esta velocidad, un aumento en la concentración del
sustrato no tiene efecto en la velocidad de la reacción.
Concentración de la enzima.- Siempre y cuando haya sustrato
disponible, un aumento en la concentración de la enzima aumenta la velocidad
enzimática hacia cierto límite.
Temperatura.- Un incremento de 10°C duplica la velocidad de
reacción, hasta ciertos límites. El calor es un factor que desnaturaliza las
proteínas por lo tanto si la temperatura se eleva demasiada, la enzima pierde
su actividad.
pH.- El
pH óptimo de la actividad enzimática es 7, excepto las enzimas del estómago
cuyo pH óptimo es ácido.
Presencia de cofactores.- Muchas enzimas dependen de los
cofactores, sean activadores o coenzimas para funcionar adecuadamente. Para las
enzimas que tienen cofactores, la concentración del cofactor debe ser igual o
mayor que la concentración de la enzima para obtener una actividad catalítica
máxima.

Zárraga, J.C.; Velázquez, I.; Rodríguez, A.;
Castells, Y. Química. México, McGraw-Hill, 2003.
PROTEÍNAS
Las funciones de las proteínas son de gran importancia aunque mucha gente piensa que sirven sólo para crear los músculos y poco más, sin embargo, las funciones de las proteínas son varias y bien diferenciadas. Las proteínas determinan la forma y la estructura de las células y dirigen casi todos los procesos vitales.
Las funciones de las proteínas son específicas de cada tipo de proteína y permiten que las células defenderse de agentes externos, mantener su integridad, controlar y regular funciones, reparar daños... Todos los tipos de proteínas realizan su función de la misma forma: Por unión selectiva a moléculasLas proteínas estructurales se unen a otras moléculas de otras proteínas y funciones que realizan incluyen la creación una estructura mayor mientras que otras proteínas se unen a moléculas diferentes: hemoglobina a oxígeno, enzimas a sus sustratos, anticuerpos a los antígenos específicos, hormonas a sus receptores específicos, reguladores de la expresión génica al ADN...
Principales funciones de las proteínas
Las funciones de las proteínas son las siguientes:
- Las proteínas tienen una función defensiva, ya que crean los anticuerpos y regulan factores contra agentes extraños o infecciones. Toxinas bacterianas, como venenos de serpientes o la del botulismo son proteínas generadas con funciones defensivas. Las mucinas protegen las mucosas y tienen efecto germicida. El fibrinógeno y la trombina contribuyen a la formación coágulos de sangre para evitar las hemorragias. Las inmunoglobulinas actúan como anticuerpos ante posibles antígenos.
- Las proteínas tienen otras funciones reguladoras puesto que de ellas están formados los siguientes compuestos: Hemoglobina, proteínas plasmáticas, hormonas, jugos digestivos, enzimas y vitaminas que son causantes de las reacciones químicas que suceden en el organismo. Algunas proteínas como la ciclina sirven para regular la división celular y otras regulan la expresión de ciertos genes.
- Las proteínas cuya función es enzimática son las más especializadas y numerosas. Actúan como biocatalizadores acelerando las reacciones químicas del metabolismo.
- Las proteínas funcionan como amortiguadores, manteniendo en diversos medios tanto el pH interno como el equilibrio osmótico. Es la conocida como función homeostática de las proteínas.
- La contracción de los músculos través de la miosina y actina es una función de las proteínas contráctiles que facilitan el movimiento de las células constituyendo las miofibrillas que son responsables de la contracción de los músculos. En la función contráctil de las proteínastambién está implicada la dineina que está relacionada con el movimiento de cilios y flagelos.
- La función de resistencia o función estructural de las proteínas también es de gran importancia ya que las proteínas forman tejidos de sostén y relleno que confieren elasticidad y resistencia a órganos y tejidos como el colágeno del tejido conjuntivo fibroso, reticulina y elastina elastina del tejido conjuntivo elástico. Con este tipo de proteínas se forma la estructura del organismo. Algunas proteínas forman estructuras celulares como las histonas, que forman parte de los cromosomas que regulan la expresión genética. Algunas glucoproteínas actuan como receptores formando parte de las membranas celulares o facilitan el transporte de sustancias.
- Si fuera necesario, las proteínas cumplen también una función energética para el organismo pudiendo aportar hasta 4 kcal. de energía por gramo. Ejemplos de la función de reserva de las proteínas son la lactoalbúmina de la leche o a ovoalbúmina de la clara de huevo, la hordeina de la cebada y la gliadina del grano de trigo constituyendo estos últimos la reserva de aminoácidos para el desarrollo del embrión.
- Las proteínas realizan funciones de transporte. Ejemplos de ello son la hemoglobina y la mioglobina, proteínas transportadoras del oxígeno en la sangre en los organismos vertebrados y en los músculos respectivamente. En los invertebrados, la función de proteínascomo la hemoglobina que transporta el oxígeno la realizas la hemocianina. Otros ejemplos deproteínas cuya función es el transporte son citocromos que transportan electrones e lipoproteínas que transportan lípidos por la sangre.
- bibliografia: http://proteinas.org.es/funciones-de-las-proteinas
Lípidos
Se llama lípidos a un conjunto de moléculas orgánicas, la mayoría biomoléculas, compuestas principalmente por carbono e hidrógeno y en menor medida oxígeno, aunque también pueden contener fósforo, azufre y nitrógeno. Tienen como característica principal ser insolubles en agua y sí en disolventes orgánicos como el benceno. A los lípidos se les llama incorrectamente grasas, cuando las grasas son sólo un tipo de lípidos, aunque el más conocido.
Los lípidos forman un grupo de sustancias de estructura química muy heterogénea, siendo la clasificación más aceptada la siguiente:
- Lípidos saponificables: Los lípidos saponificables son los lípidos que contienen ácidos grasos en su molécula y producen reacciones químicas de saponificación. A su vez los lípidos saponificables se dividen en:
- Lípidos simples: Son aquellos lípidos que sólo contienen carbono, hidrógeno y oxígeno. Estos lípidos simples se subdividen a su vez en: Acilglicéridos o grasas (cuando los acilglicéridos son sólidos se les llama grasas y cuando son líquidos a temperatura ambiente se llaman aceites) y Céridos o ceras.
- Lípidos complejos: Son los lípidos que además de contener en su molécula carbono, hidrógeno y oxígeno, también contienen otros elementos como nitrógeno, fósforo, azufre u otra biomolécula como un glúcido. A los lípidos complejos también se les llama lípidos de membrana pues son las principales moléculas que forman las membranas celulares: Fosfolípidos y Glicolípidos.
- Lípidos insaponificables: Son los lípidos que no poseen ácidos grasos en su estructura y no producen reacciones de saponificación. Entre los lípidos insaponificables encontramos a: Terpenos, Esteroides y Prostaglandinas.
¿Qué función desempeñan los lípidos en el organismo?
Principalmente las tres siguientes:
- Función de reserva energética: Los lípidos son la principal fuente de energía de los animales ya que un gramo de grasa produce 9,4 kilocalorías en las reacciones metabólicas de oxidación, mientras que las proteínas y los glúcidos sólo producen 4,1 kilocalorías por gramo.
- Función estructural: Los lípidos forman las bicapas lipídicas de las membranas celulares. Además recubren y proporcionan consistencia a los órganos y protegen mecánicamente estructuras o son aislantes térmicos como el tejido adiposo.
- Función catalizadora, hormonal o de mensajeros químicos: Los lípidos facilitan determinadas reacciones químicas y los esteroides cumplen funciones hormonales.
¿Qué tipos de grasas intervienen en la alimentación?
Recordemos, las grasas son lípidos saponificables simples, sólidos a temperatura ambiente o líquidos en cuyo caso se llaman aceites. Puede ser:
- Grasas saturadas: Son aquellas grasas que están formadas por ácidos grasos saturados (tienen todos los enlaces completos por H). Aparecen por ejemplo en el tocino, en el sebo, etcétera. Este tipo de grasas es sólido a temperatura ambiente. Son las grasas más perjudiciales para el organismo.
- Grasas insaturadas: Son grasas formadas por ácidos grasos insaturados (tienen uno o más enlaces sin completar con H) como el oleico o el palmítico. Son líquidas a temperatura ambiente y comúnmente se les conoce como aceites. Pueden ser por ejemplo el aceite de oliva o el de girasol. Son las más beneficiosas para el cuerpo humano.
Existe una regla en la dieta para el consumo de las grasas: “Las de origen vegetal son más beneficiosas que las de origen animal, y las poliinsaturadas son más beneficiosas que las saturadas”. Hay unas grasas beneficiosas para el organismo porque disminuyen el nivel del llamado “colesterol malo”. El colesterol es un lípido presente en el plasma sanguíneo y en los tejidos de los vertebrados, su exceso se asocia con enfermedades cardiovasculares. Es transportado por dos proteínas LDL (Lipoproteína de baja densidad) y HDL (Lipoproteína de alta densidad). Nos referimos a los aceites llamados “omega-3” y “omega-6”. El efecto beneficioso es debido a que con su ingesta disminuye la concentración de LDL y aumenta la de HDL (con las grasas saturadas se produce el efecto contrario). Las lipoproteínas de alta densidad (HDL) pueden retirar el colesterol de las arterias y transportarlo al hígado para su excreción. Las lipoproteínas de baja densidad (LDL) transportan el colesterol a las arterias, si su nivel es más alto que el de HDL el colesterol tenderá a fijarse en las arterias, de ahí que se les conozca como “colesterol bueno” al HDL y “colesterol malo” al LDL
bibliografia:http://recursostic.educacion.es/primaria/ludos/web/pb/al/al05.html
II CORTE
LA CELULA

La célula es la unidad
anatómica, funcional y genética de los seres vivos.
La célula es una estructura constituida por tres elementos básicos:
1.- membrana plasmática,
2.- citoplasma y
3.- material genético (ADN).
Posee la capacidad de realizar tres funciones vitales:
nutrición, relación y reproducción.
Se llaman eucariotas a las células que tienen la información genética envuelta dentro de una membrana que forman el núcleo.
Un organismo formado por células eucariotas se denomina eucarionte.
Muchos seres unicelulares tienen la información genética dispersa por su citoplasma, no tienen núcleo. A ese tipo de células se les da el nombre de procariotas.
La célula es una estructura constituida por tres elementos básicos:
1.- membrana plasmática,
2.- citoplasma y
3.- material genético (ADN).
Posee la capacidad de realizar tres funciones vitales:
nutrición, relación y reproducción.
Se llaman eucariotas a las células que tienen la información genética envuelta dentro de una membrana que forman el núcleo.
Un organismo formado por células eucariotas se denomina eucarionte.
Muchos seres unicelulares tienen la información genética dispersa por su citoplasma, no tienen núcleo. A ese tipo de células se les da el nombre de procariotas.
Esquema de una Membrana Celular

La célula está rodeada por una membrana, denominada "membrana plasmática". La membrana delimita el territorio de la célula y controla el contenido químico de la célula.
En la composición química de la membrana entran a formar parte lípidos, proteínas y glúcidosen proporciones aproximadas de 40%, 50% y 10%, respectivamente. Los lípidos forman una doble capa y las proteínas se disponen de una forma irregular y asimétrica entre ellos. Estos componentes presentan movilidad, lo que confiere a la membrana un elevado grado de fluidez.
Por el aspecto y comportamiento el modelo de membrana se denomina "modelo de mosaico fluído"
Las funciones de la membrana podrían resumirse en :
1.TRANSPORTE
El intercambio de materia entre el interior de la célula y su ambiente externo.
2.RECONOCIMIENTO Y COMUNICACIÓN
Gracias a moléculas situadas en la parte externa de la membrana, que actúan como receptoras de sustancias.
La bicapa lipídica de la membrana actúa como una barrera que separa dos medios acuosos, el medio donde vive la célula y el medio interno celular.
Las células requieren nutrientes del exterior y deben eliminar sustancias de desecho procedentes del metabolismo y mantener su medio interno estable. La membrana presenta una permeabilidad selectiva, ya que permite el paso de pequeñas moléculas, siempre que sean lipófilas, pero regula el paso de moléculas no lipófilas.
El paso a través de la membrana posee dos modalidades:
Una pasiva, sin gasto de energía, y otra activa , con consumo de energía.

1.El transporte pasivo.
Es un proceso de difusión de sustancias a través de la membrana. Se produce siempre a favor del gradiente, es decir, de donde hay más hacia el medio donde hay menos. Este tranporte puede darse por:
Difusión simple . Es el paso de pequeñas moléculas a favor del gradiente; puede realizarse a través de la bicapa lipídica o a través de canales proteicos.
1.Difusión simple a través de la bicapa (1).
Así entran moléculas lipídicas como las hormonas esteroideas, anestésicos como el éter y fármacos liposolubles. Y sustancias apolares como el oxígeno y el nitrógeno atmosférico. Algunas moléculas polares de muy pequeño tamaño, como el agua, el CO2, el etanol y la glicerina, también atraviesan la membrana por difusión simple. La difusión del
agua recibe el nombre de ósmosis
2.Difusión simple a través de canales (2).
Se realiza mediante las denominadas proteínas de canal. Así entran iones como el Na+, K+, Ca2+, Cl-. Las proteínas de canal son proteínas con un orificio o canal interno, cuya apertura está regulada, por ejemplo por ligando, como ocurre con neurotransmisores u hormonas, que se unen a una determinada región, el receptor de la proteína de canal, que sufre una transformación estructural que induce la apertura del canal.

Difusión facilitada (3).
Permite el transporte de pequeñas moléculas polares, como los aminoácidos, monosacáridos, etc, que al no poder, que al no poder atravesar la bicapa lipídica, requieren que proteínas trasmembranosas faciliten su paso. Estas proteínass reciben el nombre de proteínas transportadoras o permeasas que, al unirse a la molécula a transportar sufren un cambio en su estructura que arrastra a dicha molécula hacia el interior de la célula
.
2.El transporte activo (4).
En este proceso también actúan proteínas de membrana, pero éstas requieren energía, en forma de ATP, para transportar las moléculas al otro lado de la membrana. Se produce cuando el transporte se realiza en contra del gradiente electroquímico. Son ejemplos de transporte activo la bomba de Na/K, y la bomba de Ca.
La bomba de Na+/K+ Requiere una proteína transmembranosa que bombea Na+ hacia el exterior de la membrana y K+ hacia el interior. Esta proteína actúa contra el gradiente gracias a su actividad como ATP-asa, ya que rompe el ATP para obtener la energía necesaria para el transporte.

Por este mecanismo, se bombea 3 Na+ hacia el exterior y 2 K+ hacia el interior, con la hidrólisis acoplada de ATP. El transporte activo de Na+ y K+ tiene una gran importancia fisiológica. De hecho todas las células animales gastan más del 30% del ATP que producen ( y las células nerviosas más del 70%) para bombear estos iones. Toda la porción citoplasmática que carece de estructura y constituye la parte líquida del citoplasma, recibe el nombre de citosol por su aspecto fluido. En él se encuentran las moléculas necesarias para el mantenimiento celular.
El citoesqueleto , consiste en una serie de fibras que da forma a la célula, y conecta distintas partes celulares, como si se tratara de vías de comunicacion celulares. Es una estructura en continuo cambio. Formado por tres tipos de componentes:
1.Microtúbulos
Son filamentos largos, formados por la proteína tubulina. Son los componentes más importantes del citoesqueleto y pueden formar asociaciones estables, como:
Centriolos
Son dos pequeños cilindros localizados en el interior del centrosoma Figura 1, exclusivos de células animales. Con el microscopio electrónico se observa que la parte externa de los centriolos está formada por nueve tripletes de microtúbulos
Figura 3 . Los centriolos se cruzan formando un ángulo de 90º.

Figura 2: Cilios y flagelos
Son delgadas prolongaciones celulares móviles que presentan básicamente la misma estructura, la diferencia entre ellos es que los cilios son muchos y cortos, mientras que los flagelos son pocos y más largos.
Constan de dos partes: una externa que sobresale de la superficie de la célula, está recubierta por la membrana plasmática y contiene un esqueleto interno de microtúbulos
llamado axonema, y otra interna, que se denomina cuerpo basal del que salen las raíces ciliares que se cree participan en la coordinación del movimiento.

2.Microfilamentos
Se sitúan principalmente en la periferia celular, debajo de la membrana y están formados por hebras de la proteína actina, trenzadas en hélice, cuya estabilidad se debe a la presencia de ATP e iones de calcio. Asociados a los filamentos de miosina, son los responsables de la contracción muscular.
3.Filamentos intermedios
Formados por diversos tipos de proteínas. Son polímeros muy estables y resistentes. Especialmente abundantes en el citoplasma de las células sometidas a fuertes tensiones mecánicas (queratina, desmina ) ya que su función consiste en repartir las tensiones, que de
otro modo podrían romper la célula.
Distribución en el citoplasma de los filamentos del citoesqueleto Como se puede apreciar en los esquemas de la figura 5, los microtúbulos irradian desde una región del citoplasma denominada centro organizador de microtúbulos o centrosoma
.
.

Los microfilamentos se encuentran dispersos por todo el citoplasma; pero se concentran fundamentalmente por debajo de la membrana plasmática.
Los filamentos intermedios, se extienden por todo el citoplasma y se anclan a la membrana plasmática proporcionando a las células resistencia mecánica.

Está formado por una red de membranas que forman cisternas, sáculos y tubos aplanados. Delimita un espacio interno llamado lúmen del retículo y se halla en continuidad estructural con la membrana externa de la envoltura nuclear.
Se pueden distinguir dos tipos de retículo
:
:
1.El Retículo endoplasmático rugoso (R.E.R.)
Presenta ribosomas unidos a su membrana. En él se realiza la síntesis proteíca. Las proteínas sintetizadas por los ribosomas, pasan al lúmen del retículo y aquí maduran hasta ser exportadas a su destino definitivo.
2.El Retículo endoplasmático liso (R.E.L.)
Carece de ribosomas y está formadopor túbulos ramificados y pequeñas vesículas esféricas. En este retículo se realiza la síntesis de lípidos.
En el reticulo de las células del hígado tiene lugar la detoxificación, que consiste en modificar a una droga o metabolito insoluble en agua,en soluble en agua, para así eliminar dichas sustancias por la orina. Descubierto por C. Golgi en 1898, consiste en un conjunto de estructuras de membrana que forma parte del elaborado sistema de membranas interno de las células. Se encuentra más desarrollado cuanto mayor es la actividad celular.
La unidad básica del orgánulo es el sáculo, que consiste en una vesícula o cisterna aplanada. Cuando una serie de sáculos se apilan, forman un dictiosoma. Además, pueden observarse toda una serie de vesículas más o menos esféricas a ambos lados y entre los sáculos. El conjunto de todos los dictiosomas y vesículas constituye el aparato de Golgi.
El dictiosoma se encuentra en íntima relación con el retículo endoplásmico, lo que permite diferenciar dos caras: la cara cis, más próxima al retículo, y la cara trans, más alejada. En la cara cis se encuentran las vesículas de transición , mientras que en la cara trans, se localizan las vesículas de secreción.
El sistema de membranas comentado al principio, constituye la respuesta de las células eucariotas a la necesidad de regular sus comunicaciones con el ambiente en el trasiego de macromoléculas. Para ello, se han desarrollado dos mecanismos en los que el aparato de Golgi está involucrado.
La adquisición de sustancias se lleva a cabo por endocitosis, mecanismo que consiste en englobar sustancias con la membrana plasmática para su posterior internalización.
La expulsión de sustancias se realiza por exocitosis , mecanismo que, en último término, consiste en la fusión con la membrana celular de las vesículas que contienen la sustancia a exportar.
Estos mecanismos dan sentido funcional al aparato de Golgi:
- Maduración de las glucoproteínas provenientes del retículo.
- Intervenir en los procesos de secreción, almacenamiento , transporte y
- transferencia de glucoproteínas.
- Formación de membranas: plasmática, del retículo, nuclear..
- Formación de la pared celular vegetal.
- Intervienen también en la formación de los lisosomas.
- Es aconsejable ver este dibujo, donde se ve la relación entre el retículo endoplásmico, el aparato de Golgi y los lisosomas.

Esquema de una Célula Animal

El citoplasma
Es el espacio celular comprendido entre la membrana plasmática y la envoltura
nuclear.
Está
constituido por el citosol, el citoesqueleto y los orgánulos
celulares.
El citosol o hialoplasma
Es el medio interno del
citoplasma. Es la solución acuosa donde flotan el citoesqueleto y los ribosomas.
Está formado por un 85% de agua con un gran contenido de sustancias dispersas en
él de forma coloidal (prótidos, lípidos, glúcidos, ácidos nucleicos y
nucleótidos así como sales disueltas.
En el
citosol se producen muchas de las funciones más importantes de mantenimiento celular, como las primeras etapas de descomposición de moléculas nutritivas y la
síntesis de muchas de las grandes moléculas que constituyen la
célula. En él se
produce una ingente cantidad de reacciones metabólicas importantes: glucólisis,
gluconeogénesis, fermentación láctica, etc.
El citoesqueleto
Aparece en todas las células
eucariotas.
La composición
química es una red de fibras de proteína (microfilamentos, filamentos intermedios y microtúbulos). Sus funciones
son mantener la forma de la célula, formar pseudópodos, contraer las fibras
musculares, transportar y organizar los orgánulos celulares.

Esquema de una Célula

(1) Núcleo.
(2) Poro Nuclear
(3) Retículo endoplásmico rugoso (RER)
(4)
Retículo endoplásmico liso (SER)
(5) Ribosoma en el RER.
(6) Proteínas que
son trasportadas.
(7) Vesícula trasportadora.
(8) Aparato de Golgi
(AG).
(9) Cisterna del AG.
(10) Transmembrana de AG.
(11) Cisterna de
AG.
(12) Vesícula secretora.
(13) Membrana plasmática.
(14) Proteína
secretada.
(15) Citoplasma.
(16) Espacio extracelular.
Retículo Endoplásmático

El retículo endoplasmático es un sistema membranoso cuya estructura consiste en una red de sáculos aplanados o cisternas, sáculos globosos o vesículas y túbulos sinuosos que se extienden por todo el citoplasma y comunican con la membrana nuclear externa. Dentro de esos sacos aplanados existe un espacio llamado lúmen que almacena las sustancias. Existen dos clases de retículo endoplasmático: rugoso (con ribosomas adheridos) y liso (libres de ribosomas asociados).
Su función
primordial es la síntesis de proteínas, la síntesis de lípidos constituyentes de
membrana y la participación en procesos de detoxificación de la
célula.
Ribosomas

Los ribosomas son estructuras globulares, carentes de membrana.
Están formados químicamente
por varias proteínas asociadas a ARN ribosómico procedente del nucléolo. Pueden
encontrarse libres en el citoplasma o adheridos a las membranas del retículo
endoplasmático. Unas proteínas (riboforinas) sirven de nexo entre ambas estructuras.
Su
estructura es sencilla: dos subunidades (una mayor o otra menor) de diferente coeficiente de sedimentación.
Su
función consiste únicamente en ser el orgánulo lector del ARN mensajero, con
órdenes de ensamblar los aminoácidos que formarán la proteína.
Son orgánulos
sintetizadores de proteínas.
Mitocondrias : La central energética

Las mitocondrias son los orgánulos celulares encargados de suministrar la mayor parte de la energía necesaria para la actividad celular, actúan por tanto,como centrales energéticas de la célula y sintetizan ATP a expensas de los carburantes metabólicos (glucosa, ácidos grasos y aminoácidos).
1. Membrana
interna.
2. Membrana
externa.
3.
Cresta.
4.
Matriz.
Lisosomas

Los lisosomas
son vesículas procedentes del Aparato de Golgi que contienen enzimas digestivas como las hidrolasas ácidas
Aparato de Golgi

Se encuentra en el citoplasma de la célula.El aparato de Golgi está formado por sacos aplanados limitados por membranas.
Funciona como
una planta empaquetadora, modificando vesículas del retículo endoplasmático
rugoso.
El material nuevo de las membranas se forma en varias cisternas del
Golgi.
Dentro de las funciones que posee el Aparato de Golgi se encuentran la
glicosilación de proteínas, selección, destinación (targeting), glicosilación de
lípidos y la síntesis de polisacáridos de la matriz
extracelular.
Vacuolas

Las vacuolas son estructuras celulares, muy abundantes
en las células vegetales, contenidas en el citoplasma, de forma más o menos
esféricas u ovoideas, generadas por la propia célula al crear una membrana
cerrada que aisla un cierto volumen celular del resto del citoplasma.
Su
contenido es fluido.
Almacenan productos de nutrición o de desecho, y pueden
contener enzimas lisosómicas.
El Núcleo

El núcleo,
rodeado de una membrana propia, llamada membrana nuclear, es la parte central de
la célula, que contiene el ácido desoxirribonucleico (ADN o en inglés DNA),
donde se encuentran codificados los genes
(1) Membrana
nuclear
(2)
Ribosomas
(3) Poros
Nucleares
(4)
Nucleolo
(5)
Cromatina
(6) Núcleo
(7) Reticulo
endoplásmico
(8) Nucleoplasma
Núcleo

El núcleo es
una estructura constituida por una doble membrana, denominada envoltura nuclear
que rodea al ADN de la celula separándolo del citoplasma.
El medio interno se
denomina nucleoplasma y en el están sumergidas, más o menos condensadas, las
fibras de ADN que se llaman cromatina y corpúsculos formados por ARN conocidos
como nucleolos.
Genes

La molécula de
ADN es una hélice larga y doble, semejante a una escalera de caracol. Los
eslabones de esta cadena, que determinan el código genético de cada individuo,
se componen de pares de cuatro tipos de moléculas denominadas bases (adenina,
timina, guanina y citosina). La adenina se empareja con la timina y la guanina
con la citosina. El código genético está escrito en tripletes, de manera que
cada grupo de tres eslabones de la cadena codifica la producción de uno de los
aminoácidos, los cuales son los componentes que constituirán las
proteínas.
Cromosomas

Cada persona
posee 23 pares de cromosomas. Una de estas parejas determina el sexo con el que
se nace, adoptando el nombre de "cromosomas sexuales".
Por su forma se
identifican los cromosomas sexuales femeninos (determinan que la persona sea de
sexo femenino) como XX, y la pareja de cromosomas masculinos como XY (determinan
que la persona sea de sexo masculino).
BIBLIOGRAFIAS http://celula-uhscp.blogspot.com/
MEIOSIS

Las células de la mayoría de los organismos eucariotas poseen dos copias de cada cromosoma, uno heredado del padre y otro de la madre. El número básico de cromosomas constituye el set haploide y, al conjunto de parejas cromosómicas se lo denomina set diploide. Cada pareja cromosómica es llamada par homólogo y ambos miembros del par poseen, en general, la misma morfología.
Ambos cromosomas de cada par de homólogos llevan el mismo
tipo de información genética, es decir determinan los mismos caracteres (por
ejemplo, la formación de un pigmento). Sin embargo, cada carácter puede
presentar más de una variante (pigmento gris, marrón o verde) y ambos miembros
de cada par de homólogos puede poseer la misma o diferente variante.

Las células diploides (que simbólicamente se
representan como 2n) son aquellas que poseen dos juegos de cromosomas (los dos cromosmas de cada par de
homólogos) y las células haploides ( que se representan como n) poseen sólo un
juego de cromosomas (uno de cada par de homólogos). En la
mayoría de los eucariotas las células somáticas son diploides y se multiplican
por sucesivasdivisiones mitóticas. Sólo una población de células, generalmente
las gametas son haploides y se forman por meiosis.
Al igual que la mitosis, la meiosis está precedida por un período de
duplicación del ADN. La división
meiótica está constitutida por dos etapas, la meiosis I y la
meiosis II; en cada etapa pueden distinguirse las 4 fases (profase,
metafase, anafase y telofase).
 |
MEIOSIS I
Aunque el código genético de un ser humano se contiene dentro de 46
cromosomas, sólo la mitad de este número existe dentro
de la célula de un esperma o huevo. Si las células no tuvieran la mitad, un
huevo fertilizado contendría 92 cromosomas y sería insostenible. La meiosis, un tipo
de división celular específica a la reproducción, evita esto partiendo en dos el
número de cromosomas en una célula. La célula mostrada
aquí se dividirá dos veces, produciendo cuatro células. Cada uno de estas
células tendrá sólo el número medio de cromosomas, pero cada cromosoma contendrá la
información genética de ambos padres.
|
 |
Interphase I
Período entre las divisiones celulares.
Durante este tiempo, los cromosomas se
reproducen-replicación del ADN.
Los cromosomas se pueden observar sin
el uso de un microscopio. El par de centríolos se duplica.
Nota: el Azul indica los cromosomas del padre; el naranja indica los cromosomas de la
madre.
|
 |
Profase I
Esta es la fase más larga de la meiosis, y en ella los cromosomas homólogos intercambian fragmentos de material genético. Se divide en cinco subfases:  Leptoteno:
Comienzan a contraerse los cromosomas y se hacen
visibles Leptoteno:
Comienzan a contraerse los cromosomas y se hacen
visibles Cigoteno: Los pares de cromosomas homólogos se aparean entre sí. Durante el
apareamiento o sinapsis, cada par de homólogos forma una estructura
compleja de ADN y proteína, el
complejo sinaptonémico. Cigoteno: Los pares de cromosomas homólogos se aparean entre sí. Durante el
apareamiento o sinapsis, cada par de homólogos forma una estructura
compleja de ADN y proteína, el
complejo sinaptonémico.  Paquiteno: Los cromosomas constituyen hebras gruesas debido a que la
sinapsis es completa. Paquiteno: Los cromosomas constituyen hebras gruesas debido a que la
sinapsis es completa.Se produce el fenómeno de entrecruzamiento o crossing-over entre los cromosomas homólogos. El entrecruzamiento consiste en un intercambio de fragmentos cromosómicos entre dos cromátides homólogas (no hermanas). Suelen darse dos o tres de estos entrecruzamientos por cada par bivalente. Los nucleólos son muy conspicuos.  Diplotene: Como cada cromosoma está formado por sus dos
crmátidas hermanas y, a su vez, los dos cromosomas homólogos están apareados entre sí, en esta
etapa se observan estructuras formadas por un haz de 4 cromátidas, llamadas
tétradas. Diplotene: Como cada cromosoma está formado por sus dos
crmátidas hermanas y, a su vez, los dos cromosomas homólogos están apareados entre sí, en esta
etapa se observan estructuras formadas por un haz de 4 cromátidas, llamadas
tétradas.El apareamiento cromosómico se vuelve más laxo y pueden observarse unas estructuras llamadas quiasmas que son la manifestación citológica del entrecruzamiento. Los cromosomas homólogos comienzan a repelerse entre sí; la formación de los quiasmas es un fenómeno relevante para la correcta segregación cromosómica.  Diacinesis: Los cromosomas se condensan al máximo y forman estructuras
capaces de segregar. Diacinesis: Los cromosomas se condensan al máximo y forman estructuras
capaces de segregar.Desaparecen el núcleo y la membrana nuclear, por lo que quedan libres en el citoplasma. |
 |
Prometafase I
Como en la mitosis, la membrana nuclear y los nucléolos ya desaparecieron completamente cuando comienza esta fase. |
 |
Metafase I
Los cromosomas se hallan en el plano ecuatorial y se ha formado el huso. Los dos cromosomas homólogos se unen, cada uno a través de su centrómero, a fibras del huso que tirarán hacia polos opuestos. |
 |
Anafase I Al igual que en la mitosis los cromosomas se desplazan hacia los polos. Sin embargo, en la anafase I meiótica los que se separan (segregan) son los dos cromosomas homólogos de cada par. Cada uno de los cromosomas de cada par homólogo segrega al azar, es decir, independientemente de los cromosomas de los otros pares. |
 |
Telofase I
Esta fase varía en los diferentes organismos. A veces los cromosomas pierden su condensación y se forman las membranas nucleares alrededor de cada uno de los polos. Otras veces, los cromosomas pasan directamente a la meiosis II. Nunca ocurre una fase de síntesis, es decir de duplicación de ADN, luego de la telofase I. |
 |
Citocinesis
La telofase I finaliza con la división del citoplasma en las células hijas, proceso que se denomina citocinesis. |
 |
|
 | |
 |
Prometafase II
Las fibras del huso atan a los cromosomas. Los centriolos están ahora en los polos de la célula. Como en el prometafase de la mitosis, las fibras de ambos extremos de las células atan a cada uno de los cuatro cromosomas. |
 |
Metafase II
Los cromosomas se hallan en el plano ecuatorial y se unen al huso mitótico a través de sus centrómeros. |
 |
|
 |
|
 |
Citocinesis
La telofase II finaliza con la división del citoplasma que da lugar a las células hijas. |
MITOSIS

La división mitótica de una célula da lugar a dos células hijas, cada una de las cuales es idéntica genéticamente a la célula madre.
El mecanismo de replicación celular en todas las células, excepto en
las germinales masculinas y femeninas se conoce como mitosis.
La
mitosis es un proceso continuo que se divide en varias fases, y que
requiere de la presencia de un aparato mitótico, formado por un huso de
microtúbulos dispuestos longitudinalmente y dispuestos entre un par de
centriolos.
El intervalo
entre cada división mitótica se conoce como ciclo celular.
Durante la mitosis, la célula se divide. En esta etapa, el material genético se condensa, formando los cromosomas y éstos, que previamente se habían duplicado en la interfase, se reparten entre las dos células hijas.
Cuando el material hereditario se duplica,
en cada cromosoma ambas copias se denominan cromátides
hermanas. Durante la mitosis se separan las cromátides hermanas de cada
cromosoma de modo que el resultado final consiste en
la formación de dos células hijas genéticamente idénticas, a partir de una
célula original.
Aunque la mitosis es un proceso dinámico, se
han establecido diferentes etapas o fases que permiten estudiar más fácilmente
los eventos que ocurren en cada una de ellas.
 |
MITOSIS
Junto con el citocinesis (la división del resto de una célula), la mitosis produce una célula madre que se divide en dos células hijas. La información genética dentro de cada uno de éstas células hijas es idéntica. La célula humana contiene 46 cromosomas. Para simplificar nuestra ilustración, mostraremos sólo cuatro. |
 |
Interfase
Período entre las divisiones celulares. Durante este tiempo, los cromosomas se reproducen-replicación del ADN. Los cromosomas se pueden observar sin el uso de un microscopio. El par de centríolos se duplica. |
 |
Profase
Condensación de la cromatina y aparición de los cromosomas como estructuras visibles.* La apariencia de cada cromosoma es la de un filamento formado por dos mitades individuales (crmátides hermanas) unidas entre sí por una región cromosómica llamada centrómero o constricción primaria.* Fragmentación y desaparición de la membrana nuclear: el nucleoplasma y el citoplasma se hacen uno.* Desaparición de los nucléolos. |
 |
Prometafase
Comienzan a separarse los centriolos y se ubican en los extremos opuestos de la célula. Entre ellos se forma el uso mitótico. |
 |
Metafase
Los cromosomas se desplazan y se sitúan en la región ecuatorial de la célula.* Los cromosomas se asocian a las fibras del huso a través de los centrómeros. |
 |
Anafase
Las cromátides hermanas de cada cromosoma se separan y se desplazan hacia los polos de la célula.* El desplazamiento de cada una de las cromátides se lleva a cabo a través de las fibras del huso, al que están unidas por el centrómero que se ha dividido también. De esta forma se produce una división exacta del material genético. Hacia el final de esta fase se forman dos grupos cromosomas idénticos (lo que antes eran las cromátides) en los polos opuestos de la célula. |
 |
Telofase
Las cromátides, ahora cromosomas hijos, se hallan en ambos polos celulares. El uso mitótico ha desaparecido. Se forma la membrana nuclear alrededor del material genético. Los cromosomas se condensan y forman la cromatina. Reaparecen los nucléolos. |
 |
Citocinesis
La telofase finaliza con la división del citoplasma en las células hijas, proceso que se denomina citocinesis. |
Actividades
GAMETOGENESIS
La gametogénesis es el proceso de maduración de los gametos tanto masculinos como femeninos. En este proceso se reduce a la mitad (meiosis) el número de cromosomas.
La formación de los gametos femeninos y masculinos acontece durante la vida intraembrionaria, pero variará en la mujer y en el hombre.
La gametogénesis femenina se llama ovogénesis, y se caracteriza por que se inicia y finaliza en la vida intraembrionaria, nunca más habrá nueva formación de ovogonias, su número irá reduciéndose a lo largo de la vida hasta la menopausia, de cada oocito sólo se produce un óvulo y un corpúsculo polar no fertilizable, no existe ninguna fase final de maduración como en la espermatogénesis y todos los óvulos maduros serán portadores de un gonosoma X.
La gametogénesis masculina se llama espermatogénesis, que continua durante toda la vida del varón tras la pubertad, de una espermatogonia proceden 4 espermatozoides fecundantes y hay una fase de espermátide que debe madurar hasta la formación del espermatozoide maduro.
En gametogénesis animales multicelulares esta se lleva a cabo en los órganos especiales de las glándulas sexuales o gónadas (ovarios, testículos, glándulas sexuales hermafroditas), y se lleva a cabo en tres etapas básicas.
La primera etapa es la reproducción del sexo primordial. Gametogonia (espermatogonias y ovogonias) por medio de una serie de mitosis consecutivas
La segunda etapa es el crecimiento y la maduración de estas células, que ahora se llaman gametocitos (espermatocitos y ovocitos), que, al igual que la gametogonia, poseen una completa (por lo general diploide) de cromosomas. En este punto, el acontecimiento crucial de la gametogénesis en los animales se produce: la división de los gametocitos por medio de la meiosis, lo que provoca una reducción (reducción a la mitad) del número de cromosomas en estas células y su conversión en células haploides.
La tercera etapa es la formación de espermatozoides (o espermatozoides) y células de huevo, en esta fase los óvulos adquieren una serie de membranas embrionarias, mientras que los espermatozoides adquieren flagelos que les permiten moverse. En la meiosis muchas especies animales y la formación del huevo se completó en la hembra después de la penetración del espermatozoide en el citoplasma del oocito pero antes de la fusión de los núcleos del espermatozoide y el óvulo.
BIBLIOFRAFIA:http://www.salud.es/gametogenesis

ESPERMATOGENESIS
Se denomina gametogénesis
a la formación de células sexuales o gametas. Se realiza por medio de la
meiosis, forma de reproducción sexual de organismos superiores. La producción de
gametas se hace a partir de células germinales que en su núcleo portan el número
de cromosomas propio de cada especie, con lo cual son diploides. Luego de las
dos divisiones de la meiosis se obtienen células sexuales haploides, es decir,
con la mitad de la dotación de cromosomas. La gametogénesis se divide en
espermatogénesis, que es la producción de espermatozoides y en ovogénesis, la
producción de óvulos.
La espermatogénesis es un proceso que se lleva a cabo en
los testículos (gónadas), que son las glándulas sexuales masculinas. En su
interior se encuentran los túbulos seminíferos, pequeños conductos enrollados de
30-60 cm de longitud y 0,2 mm de diámetro cada uno. Los dos testículos contienen
alrededor de un millar de túbulos seminíferos. En el epitelio de los túbulos
asientan las células germinativas o espermatogonias y las células de Sertoli. El
corte transversal del túbulo seminífero permite distinguir las diferentes etapas
de la espermatogénesis, por ejemplo, espermatogonias en la capa basal,
espermatocitos en división meiótica o liberación de espermatozoides hacia el
lumen del túbulo.

La espermatogénesis se inicia cuando el macho alcanza la
madurez sexual. En el hombre se produce alrededor de los 12-13 años de edad.
Desde su formación en la etapa embrionaria, las células germinales permanecen en
estado latente junto a las células de Sertoli en el interior de estructuras
denominadas cordones sexuales. Un poco antes de iniciarse la pubertad, estos
cordones se hacen huecos y aumentan de tamaño, transformándose en túbulos
seminíferos.
En su interior, las células germinales producen células madres, que
a su vez originan espermatogonias A por mitosis, iniciándose la
espermatogénesis. Estas últimas, también por mitosis, generan las llamadas
espermatogonias B, las cuales se reproducen a través de múltiples divisiones
mitóticas.
Las espermatogonias poseen 46
cromosomas en la especie humana, con lo cual son células diploides. Sucesivas
transformaciones dan lugar a espermatocitos
primarios, también diploides pero de un tamaño mucho mayor. Cada
espermatocito primario sufre una primera división por meiosis (meiosis I) y
genera dos espermatocitos secundarios
haploides, es decir, con la mitad de la dotación cromosómica de la especie. A su
vez, estos pasan por la mitosis II y producen cuatro células haploides llamadas
espermátidas. A partir de este momento se
inicia la maduración de los espermatozoides mediante la diferenciación de las
espermátidas.
Este último paso se denomina espermiogénesis, que consta de las
siguientes transformaciones:
-Reducción
total del volumen citoplasmático
-Alargamiento del núcleo que se ubica en la zona anterior
(cabeza) del espermatozoide
-Ubicación de
las mitocondrias en la parte posterior de la cabeza (cuello)
-Formación de un largo flagelo a partir de los centríolos
de la espermátida

En resumen, la espermatogénesis tiene una duración de 70-75
días, lapso de tiempo necesario para la diferenciación de espermatogonias en
espermatozoides. Se inicia en la madurez sexual y se mantiene casi hasta el
final de la vida. A partir una célula germinal diploide se generan cuatro
células sexuales haploides producto de divisiones por mitosis y meiosis. Por lo
tanto, la espermatogénesis consta de tres etapas: reproducción, crecimiento y
maduración.
Espermatogénesis

Dentro de los túbulos seminíferos, rodeando a las células
germinales, se encuentran las células de
Sertoli. Cuando se llega a la pubertad, dichas células dejan de
reproducirse ni bien comienza la espermatogénesis y dan lugar a la llamada barrera hematotesticular. Esta barrera está
compuesta por células de Sertoli que se adhieren firmemente interponiéndose
entre los capilares sanguíneos y el epitelio de los túbulos seminíferos,
impidiendo así la acción inmunológica de los linfocitos.
Cabe señalar que el
proceso meiótico que se establece en la espermatogénesis genera proteínas que,
de incursionar en el torrente sanguíneo, daría lugar a la formación de
anticuerpos que atentaría contra la fertilidad de la especie. Por lo tanto, la
barrera hematotesticular cumple con la importante misión de evitar la reacción
de los linfocitos y de estimular la evolución y migración de los espermatocitos
hacia la luz tubular.
Otra función importante de las células de Sertoli es la
nutrición de las células espermatogénicas, habida cuenta de que las
espermatogonias no tienen acceso a los nutrientes que provee la sangre debido a
la barrera antes mencionada. Además, las células de Sertoli producen fructosa,
eliminan restos citoplasmáticos de las espermátidas y mantienen un medio
adecuado para la transformación de las células germinales.
Esquema de la
disposición de las células en el testículo

Microfotografía de la
disposición de las células en el testículo

Los túbulos seminíferos están rodeados de tejido conectivo
y de capilares sanguíneos. Entre estas estructuras están las células de Leydig, que adoptan formas redondeadas
o poligonales.
Las células de Leydig
Tienen por función producir testosterona,
hormona fundamental que proporciona las características sexuales masculinas en
los animales superiores. La testosterona estimula la producción de
espermatozoides, aunque en altas concentraciones puede llegar a inhibir la
espermatogénesis.
BILBLIOGRAFIA:http://hnncbiol.blogspot.com/2008/01/espermatogenesis.html
OVOGENESIS
Es la formación de células sexuales femeninas, llamadas
óvulos, que tiene lugar en los ovarios de los animales superiores.
Tal como
sucede con la espermatogénesis, la ovogénesis se lleva a cabo por medio de la
mitosis y la meiosis, partiendo de células germinales diploides. Tras dos
divisiones sucesivas, meiosis I y meiosis II, se producen cuatro células con
caracteres hereditarios recombinados y la mitad de la carga genética
(haploides). La ovogénesis y la espermatogénesis es un proceso de formación de
gametas, que en conjunto se denomina gametogénesis.
OVOGENESIS PRENATAL
En la etapa de formación embrionaria femenina, las células
germinales se reproducen por mitosis sucesivas. Al llegar a las gónadas
(ovarios) las células germinales continúan dividiéndose por mitosis donde se
producen millares de ovogonias, que son células
madres del ovario con toda la dotación genética de la especie (diploides). Las
ovogonias dan origen por división mitótica a ovocitos
primarios, también diploides. Los ovocitos primarios se rodean de células
foliculares y epiteliales planas, formando el folículo
primordial.
Alrededor del séptimo mes
de gestación, los ovocitos primarios comienzan a dividirse por meiosis I, pero
al llegar al diploteno de la profase I, se detiene la división meiótica. Este
prolongado lapso de inactividad, llamado dictiotena, culmina cuando se alcanza la pubertad,
momento en que se reinicia el proceso de ovogénesis por acción hormonal. Se
supone que las células foliculares segregan una sustancia que frena el proceso
de maduración del ovocito primario.
OVOGENESIS POSNATAL
Las niñas nacen con folículos primarios que encierran a
todos los ovocitos primarios en dictiotena, hasta que llega la madurez sexual.
En ese momento empiezan a madurar los folículos y los ovocitos primarios
aumentan de tamaño. Un poco antes de que la mujer ovule, concluye la meiosis I y
se genera un ovocito secundario haploide y el primer cuerpo polar.
Cabe señalar
que esta división no es proporcional en cuanto a volumen, ya el cuerpo polar,
que más tarde se atrofia, es muy pequeño respecto del ovocito secundario, que
obtiene casi todo su citoplasma. En la medida que exista fecundación, el ovocito
secundario reanuda la meiosis II hasta el final, formándose un ovocito haploide
maduro y un segundo y pequeño cuerpo polar que más tarde involuciona. Si no se
produce la fecundación, el ovocito secundario es eliminado durante la
menstruación.
Cerca de dos millones de ovocitos primarios se forman en
los ovarios durante la etapa embrionaria, aunque esa cantidad se reduce
aproximadamente a 400000 al nacimiento. Al llegar la pubertad, la gran mayoría
se atrofia, puesto que solo 400-500 ovocitos primarios diploides se
transformarán en ovocitos secundarios haploides a lo largo de toda la vida
reproductiva.
Con la ovulación de cada ciclo sexual de 28 días, el ovocito
secundario pasa del ovario a la trompa de Falopio, madurando de a uno por vez.
Las hormonas segregadas por la hipófisis (gonadotrofinas) ejercen su acción
sobre los ovarios.
La hormona folículo estimulante (FSH) estimula al ovocito
primario para que se convierta en secundario, mientras que la hormona
luteneizante (LH) provoca la ovulación.
Ovogénesis

FOLICULOGÉNESIS
Los folículos
ováricos son estructuras formados por un conglomerado de células granulosas que
encierran a cada ovocito en el interior del ovario. Dentro de los folículos
tiene lugar la ovogénesis. La foliculogénesis es la formación y maduración de
los folículos ováricos, a partir del folículo primordial hasta períodos
intermedios o finales. De acuerdo a la etapa de desarrollo, se distinguen
distintos tipos de folículos.
-Folículos
primordiales: se forman en la vida embrionaria y contiene una capa de
células planas epiteliales y foliculares. Rodea al ovocito primario que está en
dictiotena.
-Folículos primarios: están
constituidos por células de forma cúbica que encierran ovocitos primarios,
también en dictiotena, pero que han aumentadote tamaño.
-Folículos secundarios: tienen un diámetro cercano a
300 micras. Poseen varias capas de células granulosas que encierran a un ovocito
secundario de 90-100 micras.
-Folículos terciarios o
de De Graaf: tienen un diámetro promedio de 20 mm. Están constituidos por
varias capas de células granulosas que se van ahuecando, formando un antro que
se llena de líquido a medida que se acerca a la superficie del ovario. El
folículo terciario contiene a un ovocito secundario latente en la profase de la
mitosis I (dictiotena) que se prepara para ser expulsado hacia la trompa de
Falopio. Se calcula que se necesitan casi tres meses para que se genere un
folículo de De Graaf a partir de un folículo primordial.
Esquema del desarrollo
de los folículos ováricos

Microfotografías de un corte de
ovario



Ovogénesis y etapas de la
vida



BIBLIOGRAFIA:http://hnncbiol.blogspot.com/2008/01/ovogenesis.html
APARATO REPRODUCTOR MASCULINO
En la especie humana, el hombre produce los gametos masculinos o espermatozoides. Estas células trasmiten al nuevo ser la información genética aportada por el padre.
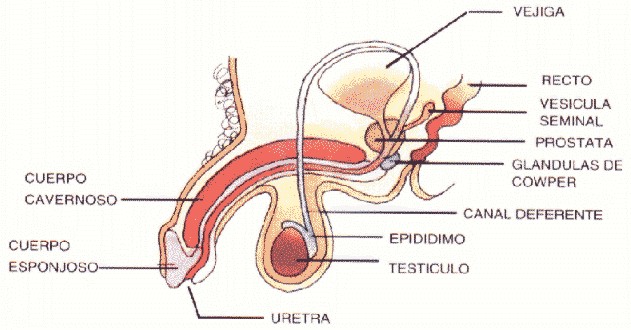 |
Las estructuras más importantes del sistema reproductor masculino son los testículos, el epidídimo, los conductos deferentes, la uretra, las vesículas seminales, la próstata y el pene.
Testículos
Son dos órganos cuya función es la producción de espermatozoides. Se encuentran suspendidos en un saco externo formado por la piel, denominado escroto. La función del escroto es mantener a los testículos en un medio más frío que el del interior de la cavidad abdominal. Para que los espermatozoides se produzcan normalmente se requiere de una temperatura 30°C menor que la temperatura corporal (37°C).
En el interior de los testículos existen unos 250 lóbulos o compartimentos, que contienen unos delgados tubos muy enrollados y apretados llamados túbulos seminíferos. Cada túbulo seminífero tiene un diámetro extremadamente pequeño y mide aproximadamente unos 80 centímetros de longitud. Son las estructuras específicas en que se producen los espermatozoides dentro del testículo.
Funcionamiento de los testículos. En el interior de los testículos y más exactamente en los túbulos seminíferos, se produce la hormona testosterona. (Ver: Hormonas sexuales) Esta hormona determina las denominadas características sexuales secundarias, y que son:
Crecimiento de la estructura del esqueleto. Es común observar en los varones, alrededor de los 12 años, un aumento importante de su estatura. Esto se debe al crecimiento de los huesos por el aumento del tejido óseo que provocan la acción de la hormona del crecimiento o somatotrofina y la testosterona. Se produce también un ensanchamiento de los hombros que dan la apariencia de un cuerpo más robusto que el de la mujer.
Los huesos del cráneo también crecen y provocan el crecimiento de la nariz, de la mandíbula y de la frente; la cara, en su conjunto, se ve más alargada en comparación con la redondeada que tenía el niño.
Desarrollo de la musculatura. El varón experimenta un aumento importante de su volumen corporal, debido al desarrollo de los músculos, más notorio en la zona pectoral y abdominal, en los bíceps y en las piernas.
Cambios en el tono de la voz. La voz cambia de un tono agudo a otro más grave, por la
maduración de la laringe y de las cuerdas vocales. En este período de cambio, es frecuente que aparezcan los conocidos "gallitos", porque el tono de la voz sube y baja involuntariamente mientras se está hablando. El desarrollo de la laringe es más notorio en los hombres porque se produce además un abultamiento en la zona delantera del cuello, que da origen a la llamada, comúnmente, "manzana de Adán", tan característica en los varones.
Cambios en la piel y en la distribución del vello.La piel del adolescente varón va engrosando, aumenta la actividad de las glándulas sebáceas y, como consecuencia, aparecen las llamadas "espinillas" y el característico acné juvenil. Estas erupciones desaparecerán cuando el cuerpo se adapte a los cambios hormonales, originados por una mayor producción de testosterona.
Aparece también abundante vellosidad en zonas del cuerpo del varón donde antes no existía: el vello facial (bigote y barba) que da la oportunidad de comenzar a afeitarse; vello en el resto del cuerpo, especialmente en las piernas, región pectoral y, más abundante, en las axilas y en la zona púbica.
La extirpación de los testículos en la pubertad hace que no se desarrollen ninguna de las características sexuales secundarias en el varón (Eunucoidismo), ya que se extrae la fuente de testosterona del individuo, además no existe la producción de espermios por lo que el individuo queda infértil.
Regulación hormonal. El funcionamiento de los testículos está controlado por una glándula cuyo nombre es adenohipófisis. Esta glándula ubicada en la base del cerebro, produce dos hormonas: la hormona folículo estimulante (HFE), que regula la producción de espermatozoides y la hormona luteinizante (HL) que controla la producción de la testosterona.
La producción de espermatozoides en el hombre está regulada por la acción de hormonas, que se mantiene constante desde la pubertad hasta la edad adulta.
Epidídimo
Es el órgano conformado por un tubo enrollado cuya longitud aproximada es de 7 centímetros. Se encuentra unido a los testículos por detrás de ellos y su función es e1 almacenar temporalmente los espermatozoides producidos en los tubos seminíferos para permitirles que adquieran movilidad. Este proceso se conoce con el nombre de capacitación, y requiere que los espermatozoides permanezcan 18 horas en el epidídimo, para completarse sólo cuando éstos ingresan al sistema reproductor femenino, donde puede ocurrir la fecundación del óvulo.
Conductos deferentes
Éstos son la prolongación del tubo contenido en el epidídimo. Su función es almacenar los gametos masculinos y transportarlos desde el testículo hasta otra porción tubular denominada uretra.
Uretra
Es un conducto que transporta los espermatozoides desde los conductos deferentes hasta el pene, para permitir su excreción.
La uretra es también el conducto por el que se elimina la orina.
Vesículas seminales
Son dos glándulas que vierten a los conductos deferentes el semen, líquido viscoso en el que flotan los espermatozoides. El semen contiene agua y nutrientes como la fructosa, un tipo de azúcar que sirve de fuente energética para posibilitar el movimiento de los espermatozoides en su camino hacia el óvulo femenino.
Próstata
Se denomina así a una glándula que segrega sustancias específicas, las cuales, al mezclarse con el semen producido por las vesículas seminales, favorece la supervivencia de los espermatozoides una vez que ingresan al sistema reproductor femenino y ocurre la fecundación del óvulo.
Pene
El pene es el órgano copulador por el cual los espermatozoides son depositados en la vagina. Está formado por un tejido esponjoso que al llenarse de sangre se separa del cuerpo en un proceso denominado erección. El pene erecto tiene la posibilidad reproductiva de introducir los espermatozoides del varón dentro del sistema reproductor femenino, función que se realiza durante el acto sexual o cópula.
En la capacidad de introducir espermas no tiene ninguna influencia el tamaño del pene.
Respecto a este punto, no existe ninguna encuesta científica ni estudio que indique algún tamaño como promedio ni menos como ideal, habiendo algunos de pocos centímetros hasta otros de quince o dieciséis. No existe tampoco ninguna correlación entre el tamaño del pene en estado de flaccidez y su estado de erección. Esto significa que un pene pequeño puede alcanzar, proporcionalmente, mayor longitud que otro de más tamaño.
APARATO REPRODUCTOR FEMENINO
La información genética que trasmite la madre a su hijo está contenida en el gameto femenino u óvulo. La producción de esta célula reproductiva pone en funcionamiento una serie de órganos sexuales que conforman el sistema reproductor femenino.
Los órganos sexuales se clasifican en internos y externos:
Los órganos internos están constituidos por:
Ovarios
Son dos órganos del tamaño de una almendra que se ubican en la cavidad abdominal de la mujer. Su función es producir un óvulo cada 28 días aproximadamente. Están situados dentro del cuerpo, en la región de la pelvis, uno a cada lado del útero.
Los ovarios producen y liberan unas hormonas denominadas estrógenos y progesterona.
Los estrógenos, producidos desde la pubertad, determinan cambios tales como: hombros angostos, voz aguda, caderas anchas, etc. Estos cambios son las características sexuales secundarias de la mujer.
La progesterona, hormona que tiene como función aumentar la cantidad de vasos sanguíneos del endometrio uterino. Cuando se libera el óvulo, una vez que ha sido fecundado, las paredes del útero están capacitadas para recibirlo y alojarlo durante su proceso de posterior desarrollo.
Trompas de Falopio
Forman un arco cerca del ovario son pequeños tubos que entran en el útero (uno derecho y otro izquierdo). Son dos conductos que se originan cerca de cada ovario y que se extienden hasta el útero. La función de las trompas, también llamadas oviductos, es conducir el óvulo desde el ovario hasta el útero. La fecundación ocurre en las trompas de Falopio.
Útero
Es un órgano musculoso y hueco del tamaño y forma de una pera invertida, y está ubicado en la parte inferior del vientre.
Lo conforman tres capas: una interna o endometrio, que cada mes se enriquece con una cantidad extra de vasos sanguíneos necesarios para la nutrición del nuevo ser; otra intermedia formada por músculos lisos; y la capa externa constituida por tejido elástico. El útero o matriz, es una cavidad que tiene cinco centímetros de longitud.
Es muscular, tiene un enorme poder de crecimiento y de contracción, pues es capaz de sacar un feto al exterior, en el momento del parto.
Vagina
Es un tubo muscular elástico que comunica el útero con el exterior. Se ubica en la pelvis menor, entre la uretra y el recto. Termina en un orificio alrededor del cual hay unos repliegues de la piel llamados labios mayores. Es un conducto que une a la vulva externa con los órganos sexuales internos. Estos se encuentran dentro de la cabida abdominal que esta situada entre los huesos de la cadera (pelvis).
Los órganos externos están formados por:
Clítoris
Pequeño órgano parecido al pene. Está provisto de terminaciones nerviosas y puede entrar en erección.
| Sistema reproductor femenino. |
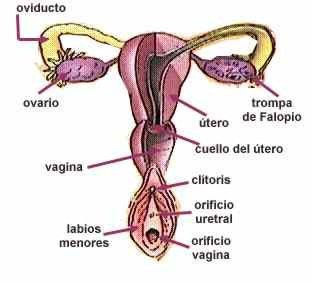
La vulva
Se puede observar al separar los muslos de la mujer. Esta rodeada de dos dobleces de piel: uno exterior, los labios mayores y uno exterior los labios menores. En la mujer púber los labios mayores tienen pelos.
Meato
Por debajo del clítoris se encuentra el meato urinario, que es el orificio de la porción final de las vías urinaria. Es el lugar donde se emite la orina al exterior.
Himen
Por la abertura de la vagina, y situado entre la entrada de ésta y el vestíbulo de ella, se encuentra en la mujer virgen una membrana no perforada llamada himen. El himen es una delgada membrana que se extiende por la abertura de la vagina.
Esta membrana tiene una o más abertura por las cuales sale el flujo menstrual y la tradición dice que en el momento de la primera penetración del órgano masculino, dicha membrana se rompe, haciendo que sangre un poco, por lo tanto era considerado una prueba de virginidad.
Actualmente se sabe que no necesariamente esta membrana se rompe en la primera relación sexual ya que puede haber membranas más elásticas que otras y además puede ser rota por otras circunstancias: utilización de tampones del diámetro no adecuado a los orificios del himen, etc.
En el sistema reproductor femenino ocurren una serie de cambios que se repiten aproximadamente cada 28 días. Las modificaciones que experimentan el útero y los ovarios constituyen el ciclo menstrual femenino.
En este ciclo de producción del gameto femenino y las hormonas sexuales femeninas se distinguen dos fases: la maduración del óvulo y secreción de estrógenos, y la ovulación y secreción de progesterona.
1. Maduración del óvulo y secreción de estrógenos
Esta fase comprende la primera mitad del ciclo, es decir dura 14 días aproximadamente. Los acontecimientos que ocurren en esta fase determinan que madure un óvulo en uno de los dos ovarios, el cual será liberado aproximadamente el día 14. Este óvulo está rodeado por células que lo nutren y protegen formando un folículo.
Paralelamente, el ovario produce y segrega estrógenos, que comenzarán a engrosar el endometrio del útero. Las paredes del útero tendrán así la capacidad de recibir al óvulo para su posterior desarrollo, en el caso de que sea fecundado.
La maduración del gameto femenino y la producción de estrógenos en los ovarios está regulada por la hormona folículo estimulante, que se origina en la adenohipófisis.
2. Ovulación y secreción de progesterona
El día 14, aproximadamente, el gameto femenino u óvulo está en condiciones de ser liberado desde el ovario.
OVULACIÓN
Es el proceso de liberación del óvulo maduro desde el ovario.
Luego de la ovulación, el folículo se transforma en una estructura del ovario llamada cuerpo lúteo, que comenzará a producir la hormona progesterona, la cual continuará los cambios iniciados por los estrógenos en el endometrio uterino.
El óvulo liberado ingresa a una de las trompas de Falopio para dirigirse hacia el útero. Si el óvulo es fecundado, se formará el cigoto, que se implantará en el endometrio uterino y comenzará su desarrollo. De lo contrario, el endometrio, con todos los vasos sanguíneos que han aumentado en cantidad y tamaño, se desintegrará produciéndose la menstruación.
MENSTRUACIÓN
Es el flujo sanguíneo liberado al exterior a través de la vagina y que contiene restos del endometrio, vasos sanguíneos y el óvulo no fecundado.
La menstruación es un proceso natural durante el cual la mujer debe procurar realizar todas sus actividades habituales. Suele durar de tres a cinco días. El primer día de la menstruación es el primer día del ciclo menstrual femenino.
CARACTERÍSTICAS SEXUALES SECUNDARIAS DE LA MUJER.
La acción hormonal provoca algunos cambios notorios en el cuerpo de la mujer joven. Estos son:
Transformación de la estructura del esqueleto. En la niña, los cambios comienzan unos dos años antes que en el hombre, es decir, alrededor de los 11 años. En ella se produce un aumento importante de la estatura, debido al crecimiento de los huesos y un ensanchamiento de las caderas. Esta última transformación es importante para la función reproductora, pues estos huesos sostendrán al feto dentro del vientre materno.
Junto con la nueva contextura de las caderas y de la pelvis se forma la cintura, que le otorga finura a la silueta femenina.
Desarrollo de las glándulas mamarias. El desarrollo de las glándulas mamarias o mamas se debe a la acción de los estrógenos. Están formadas por tejido adiposo, y por otro tejido especializado en la producción de leche, la cual se forma con las sustancias nutritivas de la dieta alimenticia, junto con el efecto de una hormona llamada prolactina que se activa después del parto.
Un conjunto de músculos presentes en el tórax son los encargados de sostener el peso de las mamas con el fin de mantenerlas en su lugar. Es recomendable realizar ciertas rutinas de ejercicios para vigorizar estos músculos.
Cambios en la piel y en la distribución del vello. Como en el varón, la acción de las hormonas sexuales provoca cambios en la textura de la piel de la mujer. Ésta se vuelve más lisa y aparecen las molestas "espinillas" por el aumento en la actividad de las glándulas sebáceas. Las alteraciones cutáneas desaparecen con el tiempo. Además, aparecen vellos, principalmente en la zona púbica y axilar.
Es propiedad: www.profesorenlinea.cl. Registro Nº 188.540
Fecundación
Los espermatozoides, atraídos por las substancias que emite el ovocito, atraviesan el cuello del útero, la cavidad uterina y se encuentran con el óvulo en una de las trompas de Falopio, cuyas vellosidades son las encargadas de transportar al óvulo hacia la cavidad uterina. La velocidad con la que el ovocito se mueve a través de la trompa es de 1,22 mm por minuto. Los espermatozoides, por su parte, se desplazan a 2-3 mm por minuto. El tiempo que tarda un espermatozoide en llegar hasta el ovocito es de unos 50 minutos
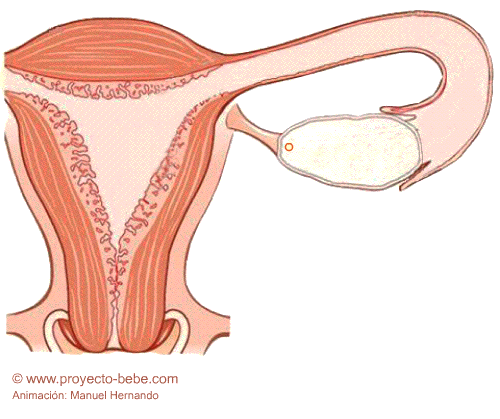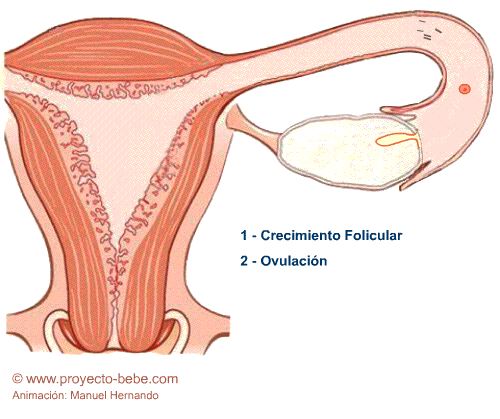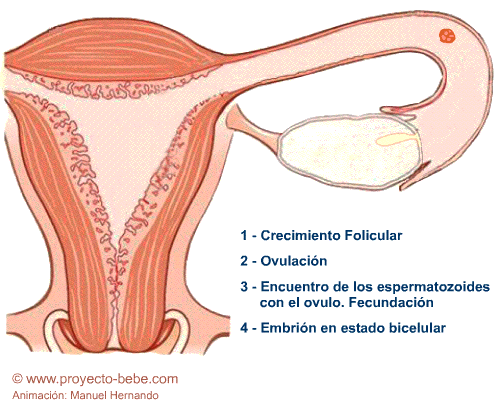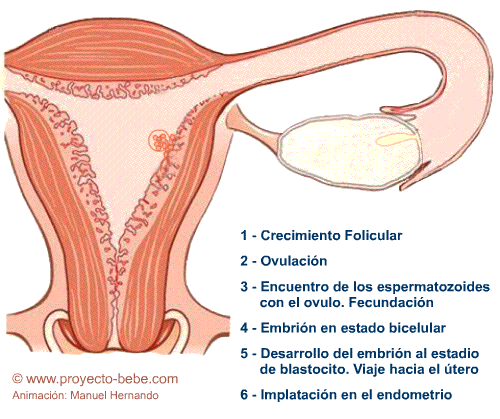
Después de que el óvulo ha sido fecundado, se transforma en una nueva célula denominada cigoto o célula huevo y comienza a descender por la trompa hacia el útero. Durante ese trayecto se inicia el proceso de la segmentación en el cual las células del cigoto se van dividiendo sucesivamente en 2, 4, 8, 16,... formando una estructura sólida similar en su forma a las moras, denominada por ello mórula.
Al quinto día post fecundación, el embrión, denominado en este estadio blastocito, ha llegado al útero. Entre dos ó cinco días más tarde se produce la implantación del embrión en la membrana uterina (llamada también endometrio), la cual fue preparada durante la fase lútea por la acción de la hormona progesterona para acoger al embrión. Muchas mujeres experimentan durante este proceso lo que se le da el nombre de sangrados de implantación. Otro fenómeno que puede acompañar el proceso de implantación es una bajada de la temperatura basal durante uno o dos días.
Una vez que el blastocito se ha implantado en el endometrio, se desarrolla el saco amniótico que albergará al embrión. El saco amniótico está lleno de líquido amniótico que amortiguará los posibles golpes que reciba.
Se inicia la formación de la placenta a su alrededor, la cual permitirá alimentar al embrión y retirar y eliminar los productos de desecho, también actuará como barrera defensiva. La comunicación entre la placenta y el embrión se realiza a través del denominado cordón umbilical, por el que pasan dos arterias y una vena.
EMBRIOLOGIA
Comienza una nueva vida - Desarrollo del embarazo paso a paso
A través de muchísimas etapas, lo que fue un pequeño óvulo fecundado se convertirá en un bebé.
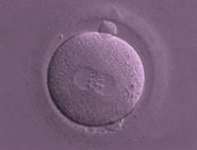 DIA 1, LA CONCEPCIÓN:
DIA 1, LA CONCEPCIÓN:
El primer paso, la fecundación del óvulo por un espermatozoide, ha tenido éxito. A partir de ahí, la información genética de el óvulo y el espermatozoide se van a reorganizar uniéndose y formando un nuevo núcleo celular compuesto por 46 cromosomas. Es el comienzo de una vida nueva y única, cuyo patrimonio genético proviene de la unión del padre y la madre. Se ha creado el embrión, al cual en este estado de desarrollo se le denomina Zigoto y mide unos 0,15 milímetros.
DIA 1,5 A 3:
El cigoto comienza su división celular. Las células del cigoto se van dividiendo sucesivamente en 2, 4, 8, hasta llegar a 16. Es entonces cuando el cigoto transforma su estructura llegando al estado de mórula. Este estado se llama así por su forma parecida a una mora. La mórula mide unos 0,2 milímetros.
DIA 4:
La mórula ha acabado su viaje a través de la trompa de Falopio y alcanza el interior del útero. Comienza una nueva transformación celular en la que las células se dividen en dos grupos. Uno de ellos comienza a formar el blastoembrión, que es lo que será el futuro embrión. El otro grupo de células van a componer lo que se denomina trofoblasto, que es la capa que va a proteger el embrión y a su vez le va a ayudar a implantarse en el endometrio.
DIA 5 A 12:
El trofoblasto segrega una serie de encimas que provocan que el endometrio sea más receptivo. El blastocito puede entonces comenzar la anidación agarrándose a las paredes del útero. El endometrio comienza a segregar la hormona hCG. Por efecto de esta hormona, el cuerpo lúteo no suspende la producción de progesterona. Es por eso que el ciclo menstrual se interrumpe y la matriz comienza a prepararse para el embarazo.
DIA 12 A 19:
Comienza la formación del saco amniótico. El embrión crece hasta alcanzar 1,5 milímetros. En estos momentos, si se hace una exploración por ecografía, se podría reconocer la existencia del embrión (implantado en el endometrio).
DIA 19 A 21:
El embrión adopta una forma como de suela de zapato. Es simétrico, posee vasos sanguíneos propios y comienza a formarse el corazón. Crece hasta 2,5 milímetros.
DIA 21 A 23:
En el embrión se ha formado el corazón y comienza a latir. Se trata de un corazón primario el cual aún no está dividido en ventrículos.
DIA 23 A 25:
El sistema central nervioso comienza a desarrollarse. El embrión mide de 3 a 4 milímetros.
DIA 25 A 27:
El embrión se muestra en forma de C. La cabeza destaca a simple vista. Ojos y orejas comienzan a formarse. El corazón comienza a desarrollar sus válvulas y tabiques. Empieza la formación de órganos digestivos.
DIA 27 A 29:
Se cumple casi el mes desde la concepción (semana 6 de embarazo). El embrión alcanza un tamaño de 6 milímetros. Comienzan a organizarse funciones vitales, tales como la respiración. Se forman la boca y la lengua. Aparecen unos pequeños estigmas que darán origen a las extremidades. Se forman las primeras células de la piel.
 DIA 29 A 41:
DIA 29 A 41:
Durante esta etapa, la cabeza y el cerebro se desarrollan rápidamente. Comienza la división cerebral de los dos hemisferios. El embrión alcanza un tamaño de 12 milímetros.
DIA 41 A 47:
Se empieza a desarrollar el primer sentido: el olfato. El centro coordinador de hormonas, la hipófisis, también ha concluido su desarrollo. En las extremidades aparecen los fundamentos de los dedos de la mano y el pié. El esqueleto, aún blando, comienza a endurecerse. Aparece la pigmentación cutánea. Los riñones producen por primera vez orina. El embrión mide unos 17 milímetros.
DIA 47 A 51:
El cerebro ha establecido el contacto con los músculos. El embrión, que ahora mide 2 centímetros puede moverse autónomamente.
DIA 51 A 55:
Los ojos están desarrollados casi completamente. Los dedos se desarrollan y aparecen unidos por una membrana como en los anfibios. El corazón ha alcanzado su desarrollo total.
DIA 55 A 57:
La mayoría de los sistemas orgánicos han concluido su formación. Los dedos están completamente desarrollados. A partir de aquí el embrión se va a denominar feto.
 SEMANA 10 A 11:
SEMANA 10 A 11:
El cerebro crece rápidamente y alcanza el desarrollo completo. En la cara comienzan a apreciarse facciones humanas. Se han formado las cuerdas vocales y el feto es capaz de emitir sonidos. Aparecen las uñas. El feto muestra reflejos, su piel es sensible. Crece alcanzando 76 mm.
SEMANA 12 A 13:
Por primera vez vas a sentir a tu feto, cuyos movimientos comienzas a notar. Podrás oír los latidos de su corazón. También se puede confirmar por ecografía el sexo.
SEMANA 14 A 17:
El feto muestra bastante actividad, moviendo a menudo su cabeza, brazos y piernas. Comienzan a formarse las huellas dactilares.
SEMANA 18 A 23:
El feto necesita dormir y comienza a buscar una posición agradable para ello. En esta etapa se le suele encontrar chupando su pulgar. También oirá la voz, la respiración y los latidos del corazón de la madre.
SEMANA 24 A 27:
Los vasos sanguíneos de los pulmones comienzan a desarrollarse para aclimatar al futuro bebé a respirar aire. La columna vertebral ha concluido su desarrollo. El feto puede abrir sus ojos, los cuales ya poseen pestañas.
SEMANA 28 A 29:
El feto mide unos 37 centímetros. El cerebro se encarga ahora regular su respiración y su temperatura corporal.
 SEMANA 30 A 31:
SEMANA 30 A 31:
Se establece la comunicación entre las células nerviosas y el cerebro. La pupila puede distinguir la luz y es capaz de contraerse. En la matriz queda poco sitio y el feto comienza a adoptar la típica posición fetal.
SEMANA 32 A 35:
Los ojos permanecen abiertos cuando el feto está despierto para cerrarse cuando duerme. Comienza a formarse el sistema inmunológico.
SEMANA 36 A 37:
El feto comienza a cambiar de posición girando su cabeza hacia abajo. Ahora puede llegar a medir 47 centímetros.

PLANIFICACION FAMILIAR
El concepto se basa en el derecho básico de la pareja proyectada como familia para poder decidir de forma responsable el tamaño de la familia en número de hijos que se quiere tener y cuanto tiempo de espaciamiento entre uno y otro, considerando las necesidades que ellos implicarán, y qué proyecto de vida aspiran a tener como familia.
La planificación familiar incentiva que se reduzcan los abortos con métodos anticonceptivos o naturales por causa de embarazos no deseados desde tempranas edades para una paternidad/maternidad responsable y exitosa como producto de una opción libre y consciente. Por supuesto esto requiere de procesos intencionales y metódicos de educación sexual tanto en el sistema educativo como familiar. Ya que no todos podemos hacerlo, porque para planificar hay que tener conocimientos de cómo hacerlo y además contar con los recursos para ello.
La planificación familiar incentiva que se reduzcan los abortos con métodos anticonceptivos o naturales por causa de embarazos no deseados desde tempranas edades para una paternidad/maternidad responsable y exitosa como producto de una opción libre y consciente. Por supuesto esto requiere de procesos intencionales y metódicos de educación sexual tanto en el sistema educativo como familiar. Ya que no todos podemos hacerlo, porque para planificar hay que tener conocimientos de cómo hacerlo y además contar con los recursos para ello.
Objetivos:
- Evitar embarazos no deseados.
- Regular los intervalos entre embarazos.
- Decidir el número de hijos en la familia.
- Regular el tiempo en que ocurren los nacimientos en relación con las edades de los padres.
- Facilitar los nacimientos deseados para mujeres con problemas de fecundidad.
- Evitar el embarazo cuando agravaría una enfermedad peligrosa presente.
- Brindar a los portadores de enfermedades genéticas la opción o posibilidad de evitar el embarazo.
La información y el conocimiento sobre fertilidad facilitan el cumplimiento de la planificación familiar, ya que implica el entendimiento de la información básica sobre fertilidad y reproducción, siendo capaz de aplicárselo a uno mismo y capaz de discutirlo con la pareja o con un profesional sanitario. Este concepto es fundamental para entender la información y tomar decisiones sobre salud reproductiva y salud sexual.
La información puede utilizarse para:
- Orientar a las parejas, tanto en la planificación de los embarazos como en evitarlos. Esto puede ser útil para ayudar a las parejas con dificultades para concebir hijos, determinando el momento ideal para las relaciones sexuales o para decidir el momento de iniciar estudios de infertilidad.
- Ayudar a las parejas a entender cómo funciona cada método anticonceptivo (cómo interrumpe la fertilidad normal, cómo falla si es usado incorrectamente y cómo se recupera la fertilidad normal al discontinuarlo).
- Las mujeres durante la lactancia, y las que se encuentran en los años perimenopáusicos, pues deben estar bien informadas acerca de la disminución de la fertilidad en esos periodos...
- Conocer en qué medida las enfermedades de transmisión sexual pueden afectar a la fertilidad..
Brindar a los portadores de enfermedades genéticas la opción o posibilidad de evitar el embarazo.
METODOS ANTICONCEPTIVOS
| Método | Descripción | Cómo funciona | Eficacia para prevenir el embarazo | Observaciones |
| Anticonceptivos orales en combinación (la «pastilla» o «píldora») | Contiene dos hormonas (estrógeno y progestágeno) | Evita la liberación de óvulos por los ovarios (ovulación) | >99% si se usa de manera correcta y sostenida | Disminuye el riesgo de cáncer endometrial y ovárico; no debe tomarse mientras se amamanta |
| 92% como se usa comúnmente | ||||
| Pastillas de progestágeno solo «minipastilla» o «minipíldora» | Contiene únicamente progesterona (sin estrógeno) | Hace más espeso el moco del conducto del cuello uterino, lo que impide que los espermatozoides y el óvulo se junten y previene la ovulación | 99% si se usa de manera correcta y sostenida | Puede usarse mientras se amamanta; debe tomarse todos los días a la misma hora |
| Entre 90% y 97% como se usa comúnmente | ||||
| Implantes | Cilindros o cápsulas pequeños y flexibles que se colocan debajo de la piel del brazo; contienen únicamente progestágeno | El mismo mecanismo de las pastillas de progestágeno | >99% | Debe ser insertado y extraído por personal sanitario; se puede usar durante 3 a 5 años, según el tipo; las hemorragias vaginales irregulares son comunes pero no dañinas |
| Progestágeno en forma inyectable | Se inyecta por vía intramuscular cada 2 o 3 meses, según el producto | El mismo mecanismo de las pastillas de progestágeno solo | >99% si se usa de manera correcta y sostenida | Al cesar el uso, la fecundidad tarda en reaparecer (entre 1 y 4 meses); las hemorragias vaginales irregulares son comunes pero no dañinas |
| 97% como se usa comúnmente | ||||
| Inyectables mensuales o anticonceptivos inyectables en combinación | Se inyectan cada mes por vía intramuscular; contienen estrógeno y progestágeno | El mismo mecanismo de los anticonceptivos orales en combinación | >99% si se usan de manera correcta y sostenida | las hemorragias vaginales irregulares son comunes pero no dañinas |
| 97% como se usan comúnmente | ||||
| Dispositivo intrauterino (DIU): de cobre | Dispositivo plástico flexible y pequeño que contiene un asa o cubierta de cobre y se inserta en el útero | El cobre daña los espermatozoides e impide que se junten con el óvulo | >99% | Durante los primeros meses la menstruación dura más y es más copiosa, pero esto no es dañino; se puede usar también como anticonceptivo de urgencia |
| Condón masculino | Vaina o cubierta que envuelve el pene erecto | Forma una barrera que impide que los espermatozoides pasen a la vagina | 98% si se usa de manera correcta y sostenida | También protege de las infecciones de transmisión sexual, en particular la causada por el VIH |
| 85% como se usa comúnmente | ||||
| Condón femenino | Vaina o forro que se adapta holgadamente a la vagina; está hecho de un material plástico transparente, fino y suave | Forma una barrera que impide que los espermatozoides y el óvulo se junten | 90% si se usa de manera correcta y sostenida | También protege de las infecciones de transmisión sexual, en particular la causada por el VIH |
| 79% como se usa comúnmente | ||||
| Esterilización masculina (vasectomía) | Anticoncepción permanente por la cual se bloquean o cortan los tubos (conductos deferentes) que transportan los espermatozoides desde los testículos | Impide que haya espermatozoides en el semen eyaculado | >99% después de la evaluación del semen a los 3 meses | Tarda en actuar unos 3 meses debido a que quedan espermatozoides almacenados; no afecta el funcionamiento sexual del hombre; es fundamental que sea una elección voluntaria y con conocimiento de causa |
| Entre 97% y 98% si no se evalúa el semen | ||||
| Esterilización femenina (ligadura de las trompas; salpingectomía) | Anticoncepción permanente por la cual se bloquean o cortan las trompas de Falopio | Los óvulos no pueden juntarse con los espermatozoides | >99% | Es fundamental que sea una elección voluntaria y con conocimiento de causa |
| Método de la retirada (coito interrumpido) | El hombre retira el pene de la vagina y eyacula fuera de esta, con lo que el semen no tiene contacto con los genitales externos de la mujer | Evita que los espermatozoides penetren en la vagina, lo que impide la fecundación | 96% si se aplica de manera correcta y sostenida | Es uno de los métodos menos eficaces porque a menudo resulta difícil determinar el momento preciso de la retirada |
| 73% como se practica comúnmente | ||||
| Métodos que requieren el conocimiento de la fecundidad (planificación natural de la familia o abstinencia periódica) | Métodos de calendario: se vigilan los días fecundos en el ciclo menstrual; métodos que se basan en síntomas: se vigilan el moco del cuello uterino y la temperatura corporaltemperature | La pareja evita el embarazo evitando tener coito sin protección durante los días fecundos, generalmente mediante la abstinencia o empleando condones | 75% | Puede usarse para determinar los días en que la mujer es fecunda, tanto si quiere embarazarse como si pretende no hacerlo |
| Método de la amenorrea del amamantamiento | Es un método de anticoncepción temporal para las mujeres recién paridas que no han vuelto a menstruar; exige el amamantamiento exclusivo, día y noche, de una criatura menor de 6 meses | Impide que los ovarios liberen óvulos (ovulación) | 99% si se aplica de manera correcta y sostenida | Es un método temporal de planificación familiar basado en el efecto natural del amamantamiento sobre la fecundidad |
| 98% como se practica comúnmente | ||||
| Anticoncepción de urgencia (levonorgestrel, 1,5 mg) | Son pastillas de progestágeno que se toman para prevenir el embarazo hasta 5 días después de una relación sexual sin protección | Evita la ovulación | Reduce en un 60% a un 90% el riesgo de embarazo | No altera el embarazo si este ya se ha producido |
TERCER CORTE

- Gen. Unidad hereditaria que controla cada carácter en los seres vivos. A nivel molecular corresponde a una sección de ADN, que contiene información para la síntesis de una cadena proteínica.
- Alelo. Cada una de las alternativas que puede tener un gen de un carácter. Por ejemplo el gen que regula el color de la semilla del guisante , presenta dos alelos, uno que determina color verde y otro que determina color amarillo. Por regla general se conocen varias formas alélicas de cada gen ; el alelo más extendido de una población se denomina "alelo normal o salvaje", mientras que los otros más escasos, se conocen como "alelos mutados".
- Carácter cualitativo. Es aquel que presenta dos alternativas claras, fáciles de observar: blanco-rojo; liso-rugoso; alas largas-alas cortas; etc. Estos caracteres están regulados por un único gen que presenta dos formas alélicas ( excepto en el caso de las series de alelos múltiples). Por ejemplo, el carácter color de la piel del guisante está regulado por un gen cuyas formas alélicas se pueden representar por dos letras, una mayúscula (A) y otra minúscula (a).
- Carácter cuantitativo. El que tiene diferentes graduaciones entre dos valores extremos. Por ejemplo la variación de estaturas, el color de la piel; la complexión física. Estos caracteres dependen de la acción acumulativa de muchos genes, cada uno de los cuales produce un efecto pequeño. En la expresión de estos caracteres influyen mucho los factores ambientales.
- Genotipo.Es el conjunto de genes que contiene un organismo heredado de sus progenitores. En organismos diploides, la mitad de los genes se heredan del padre y la otra mitad de la madre.
- Fenotipo. Es la manifestación externa del genotipo, es decir, la suma de los caracteres observables en un individuo. El fenotipo es el resultado de la interacción entre el genotipo y el ambiente. El ambiente de un gen lo constituyen los otros genes, el citoplasma celular y el medio externo donde se desarrolla el individuo.
- Locus. Es el lugar que ocupa cada gen a lo largo de un cromosoma (el plural es loci).
- Homocigoto. Individuo que para un gen dado tiene en cada cromosoma homólogo el mismo tipo de alelo, por ejemplo, AA o aa .
- Heterocigoto. Individuo que para un gen dado tiene en cada cromosoma homólogo un alelo distinto, por ejemplo, Aa.
Los principios de Mendel
La primera ley de
Mendel, o principio de segregación establece que cada individuo lleva un par de
factores para cada característica y que los miembros del par segregan –es
decir, se separan– durante la formación de los gametos. Si los miembros del par son iguales, se
dice que el individuo es homocigota para la característica determinada por ese gen; si son diferentes, el individuo es heterocigota para esa característica. Las diferentes
formas de un mismo gen son conocidas como alelos.
La constitución genética de un organismo se
denomina genotipo. Sus características externas observables se conocen como fenotipo. Un
alelo que se expresa en el fenotipo de un individuo heterocigota, con exclusión
del otro alelo, es un alelo dominante; aquel
cuyos efectos no se observan en el fenotipo del heterocigota es un alelo
recesivo. En los cruzamientos que involucran a dos individuos heterocigotas para
el mismo gen, la relación en la progenie del fenotipo dominante con respecto al
recesivo es 3:1.
Mendel cruzó una planta de guisante pura de
semillas amarillas con una planta pura de semillas verdes, transfiriendo el
polen de las anteras de las flores de una planta a los estigmas de las flores
de otra planta. Estas plantas constituyeron la generación progenitora (P). Las
flores así polinizadas originaron vainas de guisantes que contenían solamente
semillas amarillas. Estos guisantes –que son semillas– constituyeron la generación
F1. Cuando las plantas de la F1 florecieron, las dejó autopolinizarse. Las
vainas que se originaron de las flores autopolinizadas (generación F2) contenían tanto semillas amarillas como verdes, en una relación
aproximada de 3:1, o sea aproximadamente 3/4 eran amarillas y 1/4 verdes.
Una planta de guisante homocigota para flores
púrpuras, se representa como BB en símbolos genéticos ya que el alelo para flor
púrpura es dominante (B). Esta planta BB, sólo produce gametos, ya sean
femeninos o masculinos, con el alelo para flor púrpura (B). Del mismo modo, una
planta de guisante de flores blancas es homocigota recesiva (bb) y solamente
produce gametos femeninos o masculinos con el alelo para flor blanca (b).
Finalmente, una planta heterocigota (Bb) posee flores púrpura ya que el alelo
para flor púrpura (B) es dominante sobre el alelo para flor blanca (b); esta
planta produce la mitad de los gametos con el alelo B y la otra mitad, con el
alelo (b), ya sea que se trate de gametos femeninos o masculinos.
Se muestran las generaciones F1 y F2 después
de un cruzamiento entre plantas de la generación P: una planta de guisante
homocigota dominante para flores púrpuras (BB) y una planta homocigota recesiva
para flores blancas (bb).
El fenotipo de la progenie de este
cruzamiento–la generación F1– es púrpura, pero su genotipo es Bb. La F1
heterocigota produce cuatro tipos de gametos: masculinos B, femeninos B,
masculinos b y femeninos b, en proporciones iguales. Cuando esta planta se
autopoliniza, los gametos masculinos y los femeninos, B y b, se combinan al
azar y forman, en promedio 1/4 BB (púrpura), 2/4 (o 1/2) Bb (púrpura) y 1/4 bb
(blanco). La relación genotípica subyacente 1:2:1 es la que da cuenta de la
relación fenotípica de tres dominantes (púrpura) a un recesivo (blanco),
que se expresa como 3:1. La distribución de las variantes en la F2 se muestra
en un tablero de Punnett, que recibió su nombre del genetista inglés que utilizó
este tipo de diagrama para el análisis de las características determinadas
genéticamente.

Enunciado de la ley.- A esta ley se le llama también Ley de la uniformidad de los híbridos de la primera generación (F1). , y dice que cuando se cruzan dos variedades individuos de raza pura ambos (homocigotos ) para un determinado carácter, todos los híbridos de la primera generación son iguales.
El experimento de Mendel.- Mendel llegó a esta conclusión trabajando con una variedad pura de plantas de guisantes que producían las semillas amarillas y con una variedad que producía las semillas verdes. Al hacer un cruzamiento entre estas plantas, obtenía siempre plantas con semillas amarillas.

Otros casos para la primera ley.- La primera ley de Mendel se cumple también para el caso en que un determinado gen de lugar a una herencia intermedia y no dominante, como es el caso del color de las flores del "dondiego de noche" (Mirabilis jalapa). Al cruzar las plantas de la variedad de flor blanca con plantas de la variedad de flor roja, se obtienen plantas de flores rosas. La interpretación es la misma que en el caso anterior, solamente varía la manera de expresarse los distintos alelos.

Segunda ley de Mendel
Enunciado de la ley.- A la segunda ley de Mendel también se le llama de la separación o disyunción de los alelos.
El experimento de Mendel. Mendel tomó plantas procedentes de las semillas de la primera generación (F1) del experimento anterior (figura 1) y las polinizó entre sí. Del cruce obtuvo semillas amarillas y verdes en la proporción que se indica en la figura 3. Así pues, aunque el alelo que determina la coloración verde de las semillas parecía haber desaparecido en la primera generación filial, vuelve a manifestarse en esta segunada generación.

Otros casos para la segunda ley. En el caso de los genes que presentan herencia intermedia, también se cumple el enunciado de la segunda ley. Si tomamos dos plantas de flores rosas de la primera generación filial (F1) del cruce que se observa en la figura 2 y las cruzamos entre sí, se obtienen plantas con flores blancas, rosas y rojas, en la proporción que se indica en el esquema de la figura 4.También en este caso se manifiestan los alelos para el color rojo y blanco, que permanecieron ocultos en la primera generación filial.

Retrocruzamiento
Retrocruzamiento de prueba
En el caso de los genes que manifiestan herencia dominante, no existe ninguna diferencia aparente entre los individuos heterocigóticos (Aa) y los homocigóticos (AA), pues ambos individuos presentarían un fenotipo amarillo.La prueba del retrocruzamiento, o simplemente cruzamiento prueba, sirve para diferenciar el individuo homo del heterocigótico. Consiste en cruzar el fenotipo dominante con la variedad homocigota recesiva (aa).
Si es homocigótico, toda la descendencia será igual, en este caso se cumple la primera Ley de Mendel.(figura 5).
Si es heterocigótico, en la descendencia volverá a aparecer el carácter recesivo en una proporción del 50%. (figura 6).
 |  |
Tercera ley de Mendel
Enunciado de la ley.Se conoce esta ley como la de la herencia independiente de caracteres, y hace referencia al caso de que se contemplen dos caracteres distintos. Cada uno de ellos se transmite siguiendo las leyes anteriores con independencia de la presencia del otro carácter.
El experimento de Mendel. Mendel cruzó plantas de guisantes de semilla amarilla y lisa con plantas de semilla verde y rugosa ( Homocigóticas ambas para los dos caracteres).
 (Figura 7)Las semillas obtenidas en este cruzamiento eran todas amarillas y lisas, cumpliéndose así la primera ley para cada uno de los caracteres considerados , y revelándonos también que los alelos dominantes para esos caracteres son los que determinan el color amarillo y la forma lisa.
(Figura 7)Las semillas obtenidas en este cruzamiento eran todas amarillas y lisas, cumpliéndose así la primera ley para cada uno de los caracteres considerados , y revelándonos también que los alelos dominantes para esos caracteres son los que determinan el color amarillo y la forma lisa.Las plantas obtenidas y que constituyen la F1 son dihíbridas (AaBb).Estas plantas de la F1 se cruzan entre sí, teniendo en cuenta los gametos que formarán cada una de las plantas
 y que pueden verse en la figura 8. En el cuadro de la figura 9 se ven las semillas que aparecen y en las proporciones que se indica.
y que pueden verse en la figura 8. En el cuadro de la figura 9 se ven las semillas que aparecen y en las proporciones que se indica. Se puede apreciar que los alelos de los distintos genes se transmiten con independencia unos de otros, ya que en la segunda generación filial F2 aparecen guisantes amarillos y rugosos y otros que son verdes y lisos, combinaciones que no se habían dado ni en la generación parental (P), ni en la filial primera (F1).
Asímismo, los resultados obtenidos para cada uno de los caracteres considerados por separado, responden a la segunda ley.
 |
Interpretación del experimento. Los resultados de los experimentos de la tercera ley refuerzan el concepto de que los genes son independientes entre sí, que no se mezclan ni desaparecen generación trás generación. Para esta interpretación fue providencial la elección de los caracteres, pues estos resultados no se cumplen siempre, sino solamente en el caso de que los dos caracteres a estudiar estén regulados por genes que se encuentran en distintos cromosomas. No se cumple cuando los dos genes considerados se encuentran en un mismo cromosoma, es el caso de los genes ligados
Genética Post mendeliana
A principios
del siglo XX, con el redescubrimiento del trabajo de Mendel, surgieron varios
problemas hereditarios que no podían resolverse aplicando únicamente las leyes
de la herencia, esto debido a que los conceptos mendelianos se basan en la
transmisión hereditaria de características totalmente independientes entre sí.
La difusión del mendelismo hacia la investigación hereditaria experimental y la aparición de mejores microscopios durante la primera década del siglo XX, hicieron que surgiera la Genética.
Las investigaciones
posteriores a Mendel demostraron que aun cuando los genes son heredados, como
unidades independientes, existen casos en los que la manifestación de un
carácter no depende de un solo gen, sino es el resultado de la interacción
entre varios; existen otros casos en los cuales la presencia de uno solo
determina la manifestación simultánea de varios caracteres distintos.La difusión del mendelismo hacia la investigación hereditaria experimental y la aparición de mejores microscopios durante la primera década del siglo XX, hicieron que surgiera la Genética.
- HERENCIA LIGADA AL SEXO
- La especie humana posee 46 cromosomas dispuestos en 23 pares, de esos 23 pares 22 son somáticos o autosomas (heredan caracteres no sexuales) y uno es una pareja de cromosomas sexuales (llamados también heterocromosomas o gonosomas), identificados como XX en las mujeres y como XY en los hombres.
Esta pareja de cromosomas sexuales no solo llevan los genes que determinan el sexo, sino que también llevan otros que influyen sobre ciertos caracteres hereditarios no relacionados con el sexo.
Ver: Herencia y genética
Hay caracteres que sin ser caracteres sexuales primarios (órganos genitales, gónadas) o secundarios (barba del hombre, pechos de las mujeres), solo aparecen en uno de los dos sexos, o si aparecen en los dos, en uno de ellos son mucho más frecuentes.
A estos caracteres se les denomina caracteres ligados al sexo.
Variantes en la herencia ligada al sexo
Vimos que los cromosomas sexuales constituyen un par de homólogos (XX en la mujer y XY en el hombre); sin embargo, en el par XY un segmento de cada cromosoma presenta genes particulares y exclusivos (segmento heterólogo, llamado también diferencial o no homólogo), la porción restante de los cromosomas del par XY corresponde al sector homólogo, como se grafica en el esquema siguiente:
 |
Los varones sólo llevan un representante de cada gen ubicado en el sector heterólogo del X (en tanto poseen un X) y las mujeres portan dichos genes por pares (en tanto poseen dos X). Por consiguiente, la transmisión y expresión de estos genes dependen del sexo de los individuos.
Entonces, la herencia ligada al sexo se refiere a la transmisión y expresión, en los diferentes sexos, de los genes que se encuentran en el sector no homólogo (heterólogo) del cromosoma X heredado del padre.
También podemos decir que la herencia ligada al sexo no es más que la expresión en la descendencia de los genes ubicados en aquellas regiones del cromosoma X que no tienen su correspondencia en el cromosoma Y.
En el sexo femenino, la presencia de dos cromosomas X hace que los genes contenidos en estos se comporten como si se encontraran en autosomas, con normalidad.
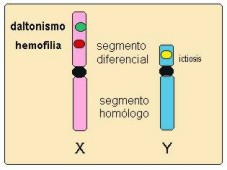 |
| Daltonismo y hemofilia, dos anomalías recesivas ubicadas en el segmento diferencial del cromosoma X. |
Si, en cambio, la mujer es heterocigota para ese carácter, igual se manifestará el alelo dominante. Es decir, se trata de un modelo de herencia clásico y normal, comparable a los mencionados en las leyes de Mendel.
El caso del hombre es radicalmente distinto. Si los genes se encuentran en la zona del cromosoma X que tiene su parte correspondiente (homóloga) en el Y, actúan como en el caso anterior.
De hecho, se han encontrado algunos alelos en los sectores homólogos de los cromosomas XY (llamados genes pseudoautosómicos) y dadas las características de estas regiones especiales de los cromosomas sexuales, los trastornos producidos a causa de estos genes se heredarán de acuerdo a las leyes genéticas que rigen la herencia autosómica (o sea, funcionan igual que los autosómicos).
Pero es mucho más frecuente que los genes estén en una parte del cromosoma X que no tenga correspondencia en el Y. Si esto sucede, los alelos se manifestarán siempre, ya sean dominantes o recesivos.
Por tanto, debemos recordar que los genes ligados a los cromosomas sexuales pueden ser tanto recesivos como dominantes.
Herencia recesiva ligada al sexo
En los hombres, los cromosomas X e Y se pueden aparear durante la meiosis (en la Profase I) por sus fragmentos homólogos y por tanto se pueden producir entrecruzamientos en esta zona.Herencia ligada al cromosoma Y o herencia holándrica
Todos los genes que se encuentran en el segmento diferencial o no homólogo del cromosoma Y son heredados únicamente por los hijos varones y se manifestarán en todos los hombres que los lleven y sólo en los hombres, independientemente de que sean dominantes o recesivos.
La transmisión de los genes situados en el segmento no homólogo del cromosoma Y es la herencia holándrica..
Holándrico:
(Del griego olos, todo, y alter, hombre). Dícese de la transmisión hereditaria de una tara o de una enfermedad que se realiza de un padre a todos sus hijos, quedando indemnes las hijas. Está ligada a genes situados en el segmento no homólogo del cromosoma sexual Y constituyéndose en una variedad de herencia ligada al sexo.
 |
| Thomas Hunt Morgan (25 de septiembre 1866 - 4 de diciembre 1945), genetista estadounidense, quien concluyó que algunos caracteres se heredan ligados al sexo. |
La herencia ligada al cromosoma X quiere decir que el gen que causa el rasgo o el trastorno se localiza en el cromosoma X .
Cabe recordar que las mujeres poseen dos cromosomas X mientras que los hombres poseen un cromosoma X y un cromosoma Y. Los genes del cromosoma X pueden ser recesivos o dominantes, y su expresión en las mujeres y en los hombres no es la misma debido a que los genes del cromosoma Y no van apareados exactamente con los genes del X.
Los genes recesivos ligados al cromosoma X se expresan en las mujeres únicamente si existen dos copias del gen (una en cada cromosoma X). Sin embargo, en los varones sólo debe haber una copia de un gen recesivo ligado al cromosoma X para que el rasgo o el trastorno se exprese.
Por ejemplo, una mujer puede ser portadora de un gen recesivo en uno de sus cromosomas X sin saberlo y transmitírselo a su hijo, que expresará el rasgo o el trastorno.
Entre los ejemplos de trastosnos recesivos ligados al cromosoma X se destacan los casos del daltonismo y la hemofilia, enfermedades provocadas por un gen recesivo situado precisamente en el segmento diferencial del cromosoma X.
Recalcamos que, debido a su ubicación, para que una mujer padezca la enfermedad debe ser homocigota recesiva (tener el gen recesivo en ambos cromosomas X), mientras que en los hombres basta con que el gen recesivo se encuentre en el único cromosoma X que tienen.
Daltonismo
Esta enfermedad, determinada por un gen recesivo del cromosoma X, es una anomalía que consiste en la incapacidad de distinguir los colores rojo y verde. Se suele llamar también ceguera para los colores, y hay muchos tipos.La enfermedad fue descrita por una persona afectada, el químico inglés John Dalton, en 1794. El nombre de esta alteración hace referencia, precisamente, a este científico.
Como ya dijimos, el gen responsable de la enfermedad es recesivo y su presencia origina el daltonismo en el hombre, mientras que la mujer que lo posee es portadora y no lo manifiesta. Para que una mujer sea daltónica es necesario que tenga genes del daltonismo en los dos cromosomas X (homocigota) , lo cual es bastante poco frecuente.
La agudeza visual (la capacidad de ver) del daltónico es normal. No existen grandes complicaciones; sin embargo, los individuos afectados pueden no ser considerados para ciertos empleos relacionados con el transporte o las Fuerzas Armadas en donde es necesario el reconocimiento de colores.
Este trastorno es 16 veces más frecuente en los hombres que en las mujeres, debido a que el gen se localiza en el cromosoma X (que es uno solo en el hombre y son dos en las mujeres; por tanto; la mujer tiene mayores posibilidades de tener el gen dominante para la visión normal en uno de los cromosomas X).
Recordemos que dominante es el alelo que se expresa y se designa con mayúscula.
Recordemos, además, que el alelo recesivo sólo se expresa cuando no está presente el dominante y se designa con minúscula.
Si caracterizamos con una letra N mayúscula (normal) la condición del cromosoma que lleva el el gen dominante que caracteriza la visión normal (sin daltonismo en nuestro caso), y con una d minúscula la tenencia del cromosoma con el el gen recesivo para el daltonismo, las posibles combinaciones que se pueden dar son:
(Recordemos que el cromosoma Y es más corto que el X y por ende es vacío en la parte donde se halla el gen que determina la visión normal).
1. Madre normal (XNXN) y padre normal (XNY):
| XN | XN | |
| XN | XNXN | XNXN |
| Y | XNY | XNY |
Ninguno de sus hijos (hombres y mujeres) será daltónico ni portador.
2. Madre normal (XNXN) y padre daltónico (XdY):
| XN | XN | |
| Xd | XdXN | XdXN |
| Y | XNY | XNY |
Todas las hijas portadoras (100 por ciento) y todos los hijos hijos normales (100 por ciento).
La combinación descrita se grafica en la imagen siguiente, donde el gen recesivo del daltonismo se identifica con la equis en rojo (X):
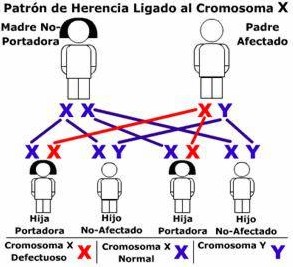 |
| Solo hijas portadoras, hijos no afectados. |
3. Madre portadora y padre normal:
| Xd | XN | |
| XN | XNXd | XNXN |
| Y | XdY | XNY |
El 50 por ciento de sus hijas seran portadoras y el 50 por ciento de sus hijos seran daltónicos.
La combinación descrita se grafica en la imagen siguiente, donde el gen recesivo del daltonismo se identifica con la equis en rojo (X):
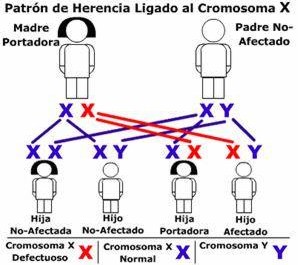 |
| Hijas e hijos, unos sanos, otros portadores. |
4. Madre portadora y padre daltónico:
| Xd | XN | |
| Xd | XdXd | XdXN |
| Y | XdY | XNY |
El 50 por ciento de hijas portadoras, 50 por ciento hijas daltónicas, 50 por ciento hijos daltónicos, 50 por ciento hijos normales.
5. Madre daltónica y padre normal:
| Xd | Xd | |
| XN | XNXd | XNXd |
| Y | XdY | XdY |
Todas las hijas portadoras y todos los hijos daltónicos.
6. Madre daltónica y padre daltónico:
| Xd | Xd | |
| Xd | XdXd | XdXd |
| Y | XdY | XdY |
Todos los hijos (hombres y mujeres) daltónicos.
Estadísticamente, lo más habitual es la madre portadora con un padre normal.
Hemofilia A
La hemofilia A es un trastorno en el cual la sangre no coagula adecuadamente debido a una insuficiencia del factor de coagulación llamado Factor VIII. El resultado es un sangrado abundante anormal que no se detiene, aun en el caso de una cortadura pequeña.A las personas con hemofilia A les aparecen moretones con facilidad y pueden tener hemorragias internas dentro de las articulaciones y los músculos.
La hemofilia A ocurre en uno de cada 10.000 varones recién nacidos.
Existe un tratamiento mediante la infusión del Factor VIII (transfusión de sangre).
Las mujeres portadoras del gen pueden mostrar signos leves de la insuficiencia del Factor VIII como los moretones que aparecen con facilidad o las hemorragias que tardan más de lo normal en detenerse luego de una cortadura. Sin embargo, no todas las mujeres portadoras presentan estos síntomas.
Se cree que un tercio de todos los casos son nuevas mutaciones en la familia (no heredadas de la madre).
Herencia dominante ligada al sexo
La herencia dominante ligada al sexo tiene lugar cuando un gen anormal de uno de los padres es capaz de causar la enfermedad, aunque pueda haber un gen compatible proveniente del otro padre que sea normal. El gen anormal domina el resultado del par de genes.Aunque la gran mayoría de las enfermedades que en la especie humana son transmitidas por genes ligados al sexo se deben, como ya vimos, a un gen recesivo, también hay ejemplos en los cuales el gen para alguna de las enfermedades es dominante, aunque los genes dominantes ligados al cromosoma X son muy poco frecuentes.
Se trata de un tipo de herencia que se caracteriza por que los varones afectados transmiten el carácter a todas sus hijas y a ninguno de sus hijos (ya que estos no reciben el cromosoma X sino que el Y, que no lleva el gen. Las mujeres afectadas, en tanto, lo transmiten a la mitad de sus hijos y a la mitad de sus hijas.
A modo de ejemplo, si hay cuatro hijos (dos varones y dos mujeres) y la madre está afectada (un X anormal y tiene la enfermedad), pero el padre no, la expectativa estadística es de:
Dos hijos (una mujer y un varón) con la enfermedad
Dos hijos (una mujer y un varón) sin la enfermedad
| XE | Xs | |
| Xs | XEXs | XsXs |
| Y | XEY | XsY |
Para mayor comprensión identificamos al gen o elelo dominante con una E mayúscula (Enfermedad) y sano o recesivo con una s minúscula (sano).
Ahora, si hay cuatro hijos (dos mujeres y dos varones) y el padre está afectado (un X anormal y tiene la enfermedad), pero la madre no, la expectativa estadística es de:
Dos niñas con la enfermedad
Dos niños sin la enfermedad
| Xs | Xs | |
| XE | XEXs | XEXs |
| Y | XsY | XsY |
Fuentes Internet:
http://www.iesbanaderos.org/html/departamentos/bio-geo/Apuntes/Bio/T14_Genet/5%20herencia%20Ligada%20sexo.htm
http://www.profes.net/rep_documentos/Pruebas_acceso_antiguas/doc6083.pdf
http://www.cocemfe.es/recursos/GuiaSalud/distrofia+muscular+duchenne/duchenne.htm
http://www.umm.edu/esp_ency/article/002050.htm
http://web.educastur.princast.es/proyectos/biogeo_ov/2BCH/b4_informacion/t410_genetica/informacion.htm
Es propiedad: www.profesorenlinea.cl. Registro Nº 188.540
Dominancia Incompleta
Carl Correns, uno de los redescubridores de Mendel cruzó
experimentalmenteflores rojas con blancas, observando en la F1 la aparición de
un nuevo fenotipo intermedio entre los parentales , es decir color rosa, lo que
no coincidía con lo descripto por Mendel, ya que la F1 era uniforme pero no
igual a uno de los progenitores.

La dominancia incompleta ocurre cuando un alelo no es
completamente dominante con respecto a otro alelo, se dice que muestra una
dominancia incompleta o ausencia de dominancia. En estos casos, ocurre que el
heterocigoto presenta un fenotipo intermedio entre los individuos homocigotos
dominantes y recesivos. Dicho en criollo no se diferencian rasgos dominantes ni
recesivos.
Al no existir
dominancia entre un alelo y el otro, se prefiere usar como notación para los
alelos, las letras mayúsculas pero con diferentes subíndices (R1 y
R2). De esta forma no se indica si uno domina sobre el otro como
usando mayúsculas y minúsculas. R1 R1 es el genotipo que
expresa fenotipo rojo, R1 R2 rosa y el R2
R2 de fenotipo blanco.

GENETICA
HUMANA
La Genética Humana describe el estudio de la
herencia al igual que ocurre en los seres humanos. La
Genética Humana abarca una variedad de campos incluidos: la genética clásica,
citogenética,
genética molecular, Biología molecular, genómica,
genética de poblaciones, genética del desarrollo, Genética médica y
el asesoramiento genético. El estudio de la genética humana puede ser útil ya que
puede responder preguntas acerca de la naturaleza humana, comprender el
desarrollo eficaz para el tratamiento de enfermedades y la genética de la vida
humana. Este artículo describe sólo características básicas de la genética
humana.

Un cariotipo es una herramienta muy útil en citogenética. Un cariotipo es la imagen de todos los cromosomas en la etapa de metafase organizado en función de la longitud y la posición del centrómero. Un cariotipo puede ser útil también en genética clínica, debido a su capacidad para diagnosticar trastornos genéticos. En un cariotipo normal, la aneuploidía puede ser detectada con claridad por la posibilidad de observar cualquier cromosoma faltante o adicional. El g-banding del cariotipo puede ser utilizado para detectar deleciones, inserciones, duplicaciones, inversiones y translocaciones. EL g-banding manchará los cromosomas con cintas claras y oscuras diferentes para cada cromosoma. La FISH, [[fluorescencia de hibridación in situ]], se puede utilizar para observar deleciones, inserciones y translocaciones. La FISH (Hibridación fluorescente in situ, por sus siglas en Inglés:"fluorescent in situ hybridization") utiliza sondas fluorescentes que se unen a secuencias específicas de los cromosomas que hará que éstos fluorezcan un único color.

Los rasgos autosómicos se asocian con un único gen en un autosoma
(cromosoma no sexual). Se les llama "dominante" porque un solo ejemplar heredado
de cualquiera de los padres es suficiente para causar la aparición de este
rasgo. A menudo, esto significa que uno de los padres también debe tener la
misma característica, a menos que ésta haya aparecido debido a una nueva
mutación. Ejemplos de
El carácter autosómico recesivo es un patrón de herencia de un rasgo,
enfermedad o trastorno que se transmite a través de las familias. Para que un
rasgo o enfermedad recesiva se manifieste, dos copias del gen (o los genes)
responsable de la aparición de ese rasgo o desorden tienen que estar presentes
en el genoma del individuo. Es decir, debe heredarse un cromosoma con el gen
portador de esa característica tanto de la madre como del padre, dando como
resultado un genotipo con dos copias del gen responsable de la aparición del
rasgo. Se denomina herencia autosómica porque el gen se encuentra en un
cromosoma autosómico: un cromosoma no sexual. Debido al hecho de que se
necesitan dos copias de un gen para expresar la característica, muchas personas
pueden, sin saberlo, ser portadores de una enfermedad. De un aspecto evolutivo,
una enfermedad o rasgo recesivo puede permanecer oculto durante varias
generaciones antes de mostrar el fenotipo. Ejemplos de trastornos autosómica
recesiva son

Cariotipo
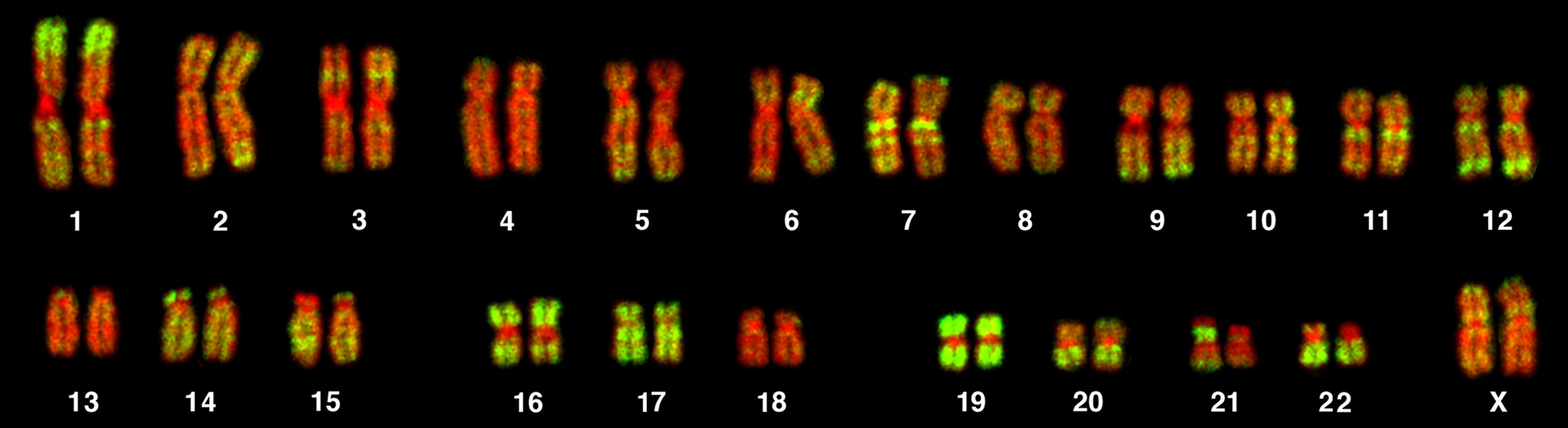
El cariotipo es el patrón cromosómico de una especie expresado a
través de un código, establecido por convenio, que describe las características
de sus cromosomas. Debido a que en el ámbito de la clínica suelen ir ligados,
el concepto de cariotipo se usa con frecuencia para referirse a un cariograma,
el cual es un esquema, foto o dibujo de los cromosomas de una célula metafásica ordenados de acuerdo a su
morfología (metacéntricos, submetacéntricos, telocéntricos, subtelocéntricos y
acrocéntricos) y tamaño, que están caracterizados y representan a todos los
individuos de una especie. El cariotipo es característico de cada especie, al
igual que el número de cromosomas; el ser humano tiene 46 cromosomas (23 pares
porque somos diploides o 2n) en el núcleo de cada célula,1 organizados en 22 pares autosómicos y 1 par sexual (hombre XY y
mujer XX).Cada brazo ha sido dividido en zonas y cada zona, a su vez, en bandas
e incluso las bandas en sub-bandas, gracias a las técnicas de marcado. No
obstante puede darse el caso, en humanos, de que existan otros patrones en los
cariotipos, a lo cual se le conoce como aberración cromosómica.
Los cromosomas se clasifican en 7 grupos, de la A a la G, atendiendo a
su longitud relativa y a la posición del centrómero, que define su morfología.
De esta manera, el cariotipo humano queda formado así:
- Grupo A: Se
encuentran los pares cromosómicos 1, 2 y 3. Se caracterizan por ser
cromosomas muy grandes, casi metacéntricos. En concreto, 1 y 3
metacéntricos; 2 submetacéntrico.
- Grupo B: Se
encuentran los pares cromosómicos 4 y 5. Se trata de cromosomas grandes y submetacéntricos
(con dos brazos muy diferentes en tamaño).
- Grupo C: Se
encuentran los pares cromosómicos 6, 7, 8, 9, 10, 11, 12, X. Son
cromosomas medianos submetacéntricos.
- Grupo D: Se
encuentran los pares cromosómicos 13, 14 y 15. Se caracterizan por ser
cromosomas medianos acrocéntricos con satélites.
- Grupo E: Se
encuentran los pares cromosómicos 16, 17 y 18. Son cromosomas pequeños,
metacéntrico el 16 y submetacéntricos 17 y 18.
- Grupo F: Se
encuentran los pares cromosómicos 19 y 20. Se trata de cromosomas pequeños
y metacéntricos.
- Grupo G: Se
encuentran los pares cromosómicos 21, 22, Y. Se caracterizan por ser
cromosomas pequeños y acrocéntricos (21 y 22 con satélites).
Mediante el cariotipado se pueden analizar anomalías numéricas y
estructurales, cosa que sería muy difícil de observar mediante genética
mendeliana.
Requisitos para el estudio del
cariotipo
Ante todo, deben guardarse las máximas condiciones de esterilidad.
Además, debe cumplirse lo siguiente:
- Las
células deben encontrarse en división. Para ello, se hará la incubación de
la muestra en presencia de productos inductores de la mitosis (mitógenos),
como es el caso de fitohemoaglutinina.
- Las
células deben pararse en prometafase, empleando colchicina, la cual
interfiere en la polimerización de los microtúbulos del huso mitótico.
- Con el
objetivo de conseguir una buena separación cromosómica, las células deben
someterse a choque osmótico. Para ello, se emplea un medio hipotónico
(0,075M KCl), que ocasiona el aumento de volumen de las células.
- Las
células tienen que ser fijadas.
- Hay que
proceder a la tinción de los cromosomas para que sean identificables.
Tinción
El estudio de los cariotipos es posible debido a la tinción. Usualmente un colorante adecuado es aplicado después
de que las [células] hayan sido detenidas durante la división celular mediante una solución de colchicina. Para humanos los glóbulos blancos son los usados más frecuentemente porque son fácilmente inducidos a
crecer y dividirse en cultivo de tejidos.2
Algunas veces las observaciones pueden ser realizadas cuando las células
no se están dividiendo (interfase). El sexo de un neonato feto puede ser determinado por
observación de células en la interfase (ver punción amniótica y corpúsculo de Barr).
La mayoría (pero no todas) las especies tienen un cariotipo estándar. El
ser humano normalmente tiene 22 pares de cromosomas autosómicos y un par de cromosomas sexuales. El cariotipo normal para la mujer contiene dos cromosoma X denominado 46,XX, y el varón
un cromosoma X y uno Y, denominado 46 XY. Cualquier variación de este cariotipo
estándar puede llevar a anormalidades en el desarrollo.
En los laboratorios de Citogenética se utilizan varias técnicas de
bandeo cromosómico. En este sentido, destaca el método de tinción de las bandas
de quinacrina (bandas Q). Fue el primero en emplearse, requiere un
microscopio de fluorescencia, aunque su uso ya no está tan extendido como el de
las bandas de giemsa (bandas G). Para producir estas bandas G
se aplica una tinción de Giemsa tras digerir parcialmente las proteínas
cromosómicas con tripsina. Las bandas reversas (bandas R)
requieren tratamiento por calor y en ellas se invierte el patrón normal blanco
y negro que se observa en las bandas Q y G. Este método destaca
por su gran utilidad en la tinción de los extremos distales de los cromosomas.
Existen otras técnicas de tinción como las bandas C y las NOR
(región de organizadores nucleolares), tiñendo estos últimos específicamente
ciertas regiones del cromosoma. Así, las bandas C tiñen la heterocromatina
constitutiva, que se localiza normalmente cerca de los centrómeros, y la
tinción NOR marca los satélites y tallos de los cromosomas
acrocéntricos.
Las bandas de alta resolución suponen la tinción de los
cromosomas en profase o metafase precoz (prometafase) antes de alcanzar la
condensación máxima. Los cromosomas en profase y prometafase están más
elongados que los cromosomas en metafase; por este motivo, el número de bandas
observadas, para el conjunto de cromosomas, aumenta desde 300-450 hasta casi
800. Ello permite detectar anomalías menos claras, que con las bandas
convencionales no suelen apreciarse. Para obtener este tipo de bandas se
necesita añadir otro requisito para la realización del cariotipo. Se trata de
un componente utilizado en quimioterapia, el metotrexato que junto con la colchicina
se añade antes de realizar la tinción.
Método de estudio del
cariotipo
1. Toma de sangre periférica y separación de los glóbulos blancos
(linfocitos T)
2. Incubación en presencia de productos que inducen a la mitosis
(mitógenos), como la fitohemoaglutinina. Los mitógenos se adicionan a las
células para que éstas crezcan adecuadamente hasta formar una monocapa. Después
se recogen separándolas del flask mediante un rascador.
3. Detención de la mitosis en la metafase (utilizando colchicina, que
interfiere en la polimerización de los microtúbulos del huso mitótico).
Tanto el paso de adición de mitógenos como la adición de colchicina son
los pasos críticos para el estudio del cariotipo.
4. Paso por un medio hipotónico que hace que las células se hinchen
5. Depositar una gota de la preparación entre porta y cubre (sobre el
cual se hace presión para dispersar los cromosomas)
6. Fijar, teñir y fotografiar los núcleos estallados (10-15; 30 en
mosaicos). Se miran de 10 a 15 núcleos porque puede haber muchos falsos
positivos debido a que le hemos añadido mitógenos (de manera que las células se
dividen de forma rápida y precipitada) y colchicina, los cuales pueden provocar
mutaciones e irregularidades en los cromosomas. Si los 10-15 núcleos no son
iguales puede ser debido a estas sustancias o a que nos encontremos delante de
un organismo mosaico (por lo que deberíamos mirar más núcleos)
7. Actualmente existen aparatos de captación y programas de análisis que
elaboran el cariotipo automáticamente con los datos obtenidos.
Es necesario realizar un recuento de, al menos, 12-25 células en
metafase. Esto es debido a que si por ejemplo, al contar un núcleo le falta el
cromosoma 21, puede ser que sea mosaico y que el resto de células sí presenten
ese cromosoma. Otra opción, y más probable, es que sea un efecto de la adición
del mitógeno, ya que este compuesto altera el proceso normal de división
celular, favoreciendo las aneuploidías. Una última opción podría ser que los
cromosomas se solapen y al proceder al analizar el cariotipo sólo se cuente un
cromosoma cuando realmente hay dos. Por todas estas razones se deben contar un
mínimo de 12-15 células en metafase que se encuentren bastante separados en el
porta.
Cariotipo clásico
En el cariotipo clásico se suele utilizar una solución de Giemsa como tinción (específica para
los grupos fosfato del ADN) para colorear las bandas de los cromosomas
(Bandas-G), menos frecuente es el uso del colorante Quinacridina (se une a las
regiones ricas en Adenosina-Timina). Cada cromosoma tiene un patrón
característico de banda que ayuda a identificarla.
Los cromosomas se organizan de forma que el brazo corto de este quede
orientado hacia la parte superior y el brazo largo hacia la parte inferior.
Algunos cariotipos nombran a los brazos cortos p y a los largos q. Además, las
diferentes regiones y subregiones teñidas reciben designaciones numéricas según
la posición a la que se encuentren respecto a estos brazos cromosómicos. Por
ejemplo, el síndrome de Cri du Chat implica una deleción en el brazo corto del cromosoma 5. Está escrito
como 46, XX, 5p-. La región crítica para este síndrome es la deleción de 15.2,
la cual es escrita como 46,XX, del(5)(p15.2)3
Cariotipo espectral

El análisis espectral de los cariotipos (o SKY) se trata de una
tecnología de citogenética molecular que permite el estudio y visualización de
los 23 pares de cromosomas en forma simultánea. Sondas marcadas
fluorescentemente son hechas para cada cromosoma al marcar DNA específico de
cada cromosoma con diferentes fluoróforos. Debido a que hay un limitado número
de fluoróforos espectralmente distintos, un método de etiquetado combinatorio
es usado para generar muchos colores diferentes. La diferencias espectrales
generadas por el etiquetado combinatorio son capturadas y analizadas usando un
interferómetro agregado a un microscopio de fluorescencia. El programa de
procesamiento de imágenes entonces asigna un pseudocolor a cada combinación
espectralmente diferente, permitiendo la visualización de cromosomas
coloreados.4
Esta técnica es usada para identificar aberraciones estructurales
cromosómicas en células cancerígenas y otras patologías cuando el bandeo con
Giemsa u otras técnicas no son lo suficientemente precisas.
Este tipo de técnicas mejorará la identificación y diagnóstico de las
aberraciones cromosómicas en citogenética prenatal así como en células
cancerosas.
Cariotipo digital
El cariotipo digital es una técnica utilizada para cuantificar el número
de copias de ADN en una escala genómica. Se trata de secuencias de locus de ADN
específicos de todo el genoma que son aisladas y enumeradas. 5
Observaciones en cariotipo
- Los
cromosomas sufren grandes variaciones en su tamaño a lo largo del ciclo
celular, pasando de estar muy poco compactados (interfase) a estar muy
compactados (metafase).
- Diferencia
de posición del centrómero.
- Las
diferencias en el número básico de cromosomas puede ocurrir debido a
desplazamientos sucesivos que quitan todo el material genético de un
cromosoma, haciendo que este se pierda.
- Diferencias
de grado y distribución de regiones de heterocromatina. La
heterocromatina, es una forma inactiva de ADN condensada localizada sobre
todo en la periferia del núcleo que se tiñe fuertemente con las
coloraciones, tomando coloración más oscura que la cromatina.
La variación de estos cromosomas es encontrada frecuentemente:
- Entre
sexos
- Entre
gametos y el resto del cuerpo.
- Entre
los miembros de una población.
- Variación
geográfica
CROMOSOMAS METAFASICOS
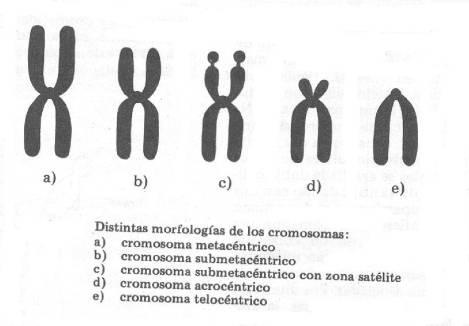

El soporte físico de la herencia
Si bién el período entre principios de siglo y la Segunda Guerra Mundial (1900 a 1940) ha sido considerado la edad de oro de la genética, los científicos aún no habían determinado que, en el ADN y no en las proteínas, se encontraba el material hereditario. Sin embargo en esa época se realizaron muchos descubrimientos genéticos y se estableció la relación entre genética y evolución.
El ADN fué aislado por Meischer en 1869 de esperma de salmón y de pús de heridas abiertas. Dado que la encontró solamente en los núcleos, Meischer denominó a este compuesto nucleína. A posteriori se lo cambió a ácido nucléico y por último a ácido desoxirribonucleico(ADN).
Robert Feulgen, en 1914, describió un método para revelar por tinción el ADN, basado en el colorante fucsina. Se encontró, utilizando este método, la presencia de ADN en el núcleo de todas las células eucariotas, específicamente en los cromosomas.
Durante los años 20, el bioquímico P.A. Levene analizó los componentes del ADN. Encontró que contenía cuatro bases nitrogenadas: citosina, timina, adenina, y guanina; el azúcar desoxirribosa; y un grupo fosfato.
El concluyó:
1) que la unidad básica (nucleótido) estaba compuesta de una base pegada a un azúcar y que el fosfato también estaba pegado al azúcar y ,
2) lamentablemente también concluyó erroneamente que las bases estaban en cantidades iguales y, que un tetranucleótido era la unidad repetitiva de la molécula.
Sin embargo queda su idéa de la estructura del nucleótido el cual es realmente la unidad fundamental (monomero) del polímero ácido nucléico.
Existen cuatro nucleótidos: uno con citosina (C), uno con guanina (G), uno con adenina (A), y uno con timina (T), y se muestran aquí en su forma "activa", como trinucleótidos, antes de entrar en la molécula de ADN, recuerde que el nucleótido tiene un solo fosfato.



http://www.whfreeman.com/life/update/.
A comienzo del siglo pasado (mejor precisemos "del año 1900" por si estan afectados por el 2YK), el estudio de la genética comienza a dar frutos: la relación entre el trabajo de Mendel y el de los biólogos celulares resultó en la teoría cromosómica de la herencia; Garrod propuso la relación entre los "errores innatos del metabolismo" y los genes. La pregunta quedó planteada: ¿que es un gen? .
La repuesta la trajo el estudio de una enfermedad infecciosa mortal la neumonía. Durante los años 20 (192... por supuesto) Frederick Griffith estudió las diferencias entre una cepa de la bacteria Streptococcus peumoniae que producía la enfermedad y otra que no la causaba. La cepa que causaba la enfermedad estaba rodeada de una cápsula ( también se la conoce como cepa S ,del ingles smooth, o sea lisa, que es el aspecto de la colonia en las placas de Petri). La otra cepa ( la R , de rugosa, que es el aspecto de la colonia en la placa de Petri) no tiene cápsula y tampoco causa neumonía. Frederick Griffith (1928) fué capaz de inducir la transformación de una cepa no patogénica Streptococcus pneumoniae EN PATOGÉNICA. Griffith postuló la existencia de un factor de transformación como responsable de este fenómeno.
Griffith inyectó las diferentes cepas de la bacteria en ratones. La cepa S mataba a los ratones mientras que la cepa R no lo hacía. Luego comprobó que la cepa S, muerta por calentamiento, no causaba neumonía cuando se la inyectaba. Sin embargo cuando combinaba la cepa S muerta por calentamiento, con la cepa R viva, e inyectaba la mezcla a los ratones ( recuerde que ningún componente individual de la mezcla mata a los ratones) los ratones contraían la neumonía y morían.
Las bacterias que se aislaban de los ratones muertos poseían cápsula y , cuando se las inyectaba, mataban otros ratones!
Hipótesis
1. La cepa S, muerta por el calor, fué reanimada o resurreció.
2. La cepa R viva fué modificada por algún "factor de transformacion" (transforming factor).
Otros experimentos mostraron que la segunda postulación era la correcta.
En los años 40 (19..), Oswald Avery, Colin MacLeod, y Maclyn McCarty revisaron el experimento de Griffith y concluyeron que el factor de transformación era el ADN. Oswald Avery demostró que el factor de transformación era el ADN repitiendo el trabajo de Griffith con el agregado de una enzima que destruía el ADN. Cuando Avery agregaba esta enzima, no observaba la transformación obtenida por Griffith . El concluyó que el material hereditario era ADN y no una proteína. Su evidencia era fuerte pero no totalmente conclusiva , para esa época el "candidato principal" para ser el material hereditario eran las proteínas.
La última palabra en la cuestión de determinar cual era el material hereditario vino de los trabajos de Max Delbruck y Salvador Luria en los 40. Los bacteriófagos son un tipo de virus que atacan a las bacterias, Delbruck y Luria trabajaron con virus que atacan a la bacteria del intestino humano Escherichia coli. Los bacteriófagos consisten en ADN con una cubierta de proteínas . Los bacteriófagos infectan una célula inyectándole su ADN. Este ADN viral "desaparece" mientras toma control de la maquinaria de la bacteria que comienza a fabricar nuevos virus. Luego de 25 minutos de haber sido inyectado la célula hospedadora estalla, liberando cientos de nuevos bacteriófagos. Como los fagos tienen solo ADN y Proteínas, eran la herramienta ideal para resolver la naturaleza del material hereditario.

En 1952 *Alfred D. Hershey y Martha Chase realizaron una serie de experimentos destinados a dilucidar si el ADN o las proteínas eran el material hereditario. Marcando el ADN y las proteínas con isótopos radioactivos el experimento demostraría cual de ellos entraba en la bacteria. Ese sería el material hereditario ( factor transformador de Griffith). Dado que el ADN contiene fósforo (P) pero no azufre (S), ellos marcaron el ADN con Fósforo-32 radioactivo. Por otra parte, las proteínas no contienen P pero si S, y por lo tanto se marcaron con Azufre-35. Hershey y Chase encontraron que el S-35 queda fuera de la célula mientras que el P-32 se lo encontraba en el interior, indicando que el ADN era el soporte físico de la herencia.




Las figuras de arriba son modificadas de: University of Arizona's Bio 181 Page.
La estructura del ADN
Erwin Chargaff analizó las base nitrogenadas del ADN en diferentes formas de vida, concluyendo que, la cantidad de purinas no siempre se encontraban en proporciones iguales a las de las pirimidinas (contrariamente a lo propuesto por Levene), la proporción era igual en todas las células de los individuos de una especie dada, pero variaba de una especie a otra.
Los experimentos de Hershey-Chase probaron que el ADN era el material genético pero, no como el ADN conformaba los genes. El ADN debía transferir información de la célula de orígen a la célula hija. Debía también contener información para replicarse a si mismo, ser químicamente estable y tener pocos cambios. Sin embargo debía ser capaz de cambios mutacionales. Sin mutaciones no existiría el proceso evolutivo.
Muchos científicos se interesaron en descifrar la estructura del ADN, entre ellos, Francis Crick, James Watson, Rosalind Franklin, y Maurice Wilkins.
Watson y Crick ( laboratorio Cavendish, Cambridge, Inglaterra) integraron todos los datos disponibles en un intento de desarrollar un modelo de la estructura del ADN. Franklin tomó fotomicrografías de difracción de rayos X de cristales de ADN, que fueron la pieza clave del rompecabezas. Los datos que se conocían por ese tiempo eran :
1) que el ADN era una molécula grande también muy larga y delgada.
2) los datos de las bases proporcionados por Chargaff (A=T y C=G; purinas/pirimidinas=k para una misma especie).
3) los datos de la difracción de los rayos-x de Franklin y Wilkins (King's College de Londres).
4) los trabajos de Linus Pauling sobre proteínas ( forma de hélice mantenida por puentes hidrógeno), quién sugirió para el ADN una estructura semejante.
El ADN es una doble hélice, con las bases dirigidas hacia el centro, perpendiculares al eje de la molécula (como los peldaños de una escalera caracol) y las unidades azúcar-fosfato a lo largo de los lados de la hélice (como las barandas de una escalera caracol). Las hebras que la conforman son complementarias (deducción realizada por Watson y Crick a partir de los datos de Chargaff, A se aparea con T y C con G, el apareamiento se mantiene debido a la acción de los puentes hidrogeno entre ambas bases). Tome nota que una purina con doble anillo siempre se aparea con una pirimidina con un solo anillo en su molécula.
Las purinas son la Adenina (A) y la Guanina (G). Durante este curso hablamos del Adenosin trifosfato (ATP) , pero en ese caso el azúcar era la ribosa, mientras que en el ADN se encuentra la desoxirribosa.
Las Pirimidinas son la Citosina (C) y la Timina (T).
Las bases son complementarias, con A en un lado de la molécula únicamente encontramos T del otro lado, lo mismo ocurre con G y C. Si conocemos la secuencia de bases de una de las hebras, conocemos su complementaria.

http://www.whfreeman.com/life/update/.
Replicación del ADN
Una vez que se comprobó que el ADN era el material hereditario y se descifró su estructura, lo que quedaba era determinar como el ADN copiaba su información y como la misma se expresaba en el fenotipo. Matthew Meselson y Franklin W. Stahl diseñaron el experimento para determinar el método de la replicación del ADN. Tres modelos de replicación era plausibles.
1.
Replicación
conservativa
Durante la cual se
produciría un ADN completamente nuevo durante la replicación.

http://www.whfreeman.com/life/update/.
2.
En
la replicación semiconservativa
se originan dos
moléculas de ADN, cada una de ellas compuesta de una hebra de el ADN original y
de una hebra complementaria nueva . En otras palabras el ADN se forma de una
hebra vieja y otra nueva. Es decir que las hebras existentes sirven de molde
complementario a las nuevas.

3.
La replicación dispersiva
implicaría
la ruptura de las hebras de origen durante la replicación que, de alguna manera
se reordenarián en una molécula con una mezcla de fragmentos nuevos y viejos en
cada hebra de ADN.

http://www.whfreeman.com/life/update/.
El experimento de Meselson-Stahl consiste en cultivar la bacteria Escherichia coli en un medio que contenga nitrógeno pesado (Nitrogeno-15 que es mas pesado que el isótopo mas común: el Nitrogeno-14, ). La primera generación de bacterias se hizo crecer en un medio que únicamente contenía Nitrogeno-15 como fuente de N. La bacteria se transfirió luego a un medio con N-14. Watson y Crick habían pronosticado que la replicación del ADN era semiconservativa, de ser así el ADN extraído de las bacterias luego de cultivarlas por una generación en N-14 tendría un peso intermedio entre el ADN extraído del medio con N-15 y el del extraído de medio con N-14, y así fue.
Los detalles del experimento que incluye un proceso de ultracentrifugación en cloruro de Cesio (CeCl2) puede encontrarse en el Curtis, pag. 316.
La replicación del ADN,que ocurre una sola vez en cada generación celular, necesita de muchos "ladrillos", enzimas, y una gran cantidad de energía en forma de ATP (recuerde que luego de la fase S del ciclo celular las células pasan a una fase G a fin de, entre otras cosas, recuperar energía para la siguiente fase de la división celular). La replicación del ADN en el ser humano a una velocidad de 50 nucleótidos por segundo, en procariotas a 500/segundo. Los nucleótidos tienen que se armados y estar disponibles en el núcleo conjuntamente con la energía para unirlos.
La iniciación de la replicación siempre acontece en un cierto grupo de nucleótidos, el origen de la replicación, requiere entre otras de las enzimas helicasas para romper los puentes hidrógeno y las topoisomerasas para aliviar la tensión y de las proteínas de unión a cadena simple para mantener separadas las cadenas abiertas.
Una vez que se abre la molécula, se forma una área conocida como "burbuja de replicación" en ella se encuentran las "horquillas de replicación" . Por acción de la la ADN polimerasa los nuevos nucleótidos entran en la horquilla y se enlazan con el nucleótido correspondiente de la cadena de origen (A con T, C con G). Los procariotas abren una sola burbuja de replicación, mientras que los eucariotas múltiples. El ADN se replica en toda su longitud por confluencia de las "burbujas".
Dado que las cadenas del ADN son antiparalelas, y que la replicación procede solo de en la dirección 5' to 3' en AMBAS cadenas, numerosos experimentos mostraron que, una cadena formará una copia continua, mientras que en la otra se formarán una série de fragmentos cortos conocidos como fragmentos de Okazaki . La cadena que se sintetiza de manera continua se conoce como cadena adelantada y la que se sintetiza en fragmentos cadena atrasada.
Para que trabaje la ADN polimerasa es necesario la presencia, en el inicio de cada nuevo fragmento, de pequeñas unidades de ARN conocidas como cebadores, a posteriori, cuando la polimerasa toca el extremo 5' de un cebador, se activan otras enzimas, que remueven los fragmentos de ARN, colocan nucleótidos de ADN en su lugar y, una ADN ligasa los une a la cadena en crecimeinto
http://www.whfreeman.com/life/update/.

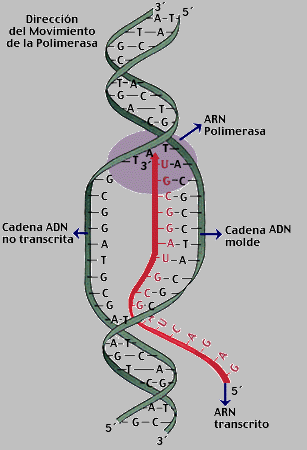


http://www.whfreeman.com/life/update/
Esquema del proceso de transcripción de ADN. Se muestra
la orientación antiparalela de las cadenas de ADN y la producción de ARN por
acción de la enzima ARN polimerasa
La transcripción del ADN
es el primer proceso de la expresión génica, mediante el cual se transfiere la información contenida en la secuencia del ADN hacia la secuencia de proteína utilizando diversos ARN como intermediarios. Durante la transcripción genética, las secuencias de ADN son copiadas a ARN mediante una enzima llamada ARN polimerasa que sintetiza un ARN mensajero que mantiene la información de la secuencia del ADN. De esta manera, la transcripción del ADN también podría llamarse síntesis del ARN mensajero.
Transcripción en eucariotas
En el caso de las eucariotas, el proceso se realiza en el núcleo, y es similar al de las procariotas, pero de mayor complejidad. Diferentes ARNp transcriben distintos tipos de genes. La ARNpII transcribe los pre-ARNm, mientras que la ARNpI y ARNpIII transcriben los ARN-ribosomales y ARNt, respectivamente. Los ARNs transcritos son modificados posteriormente. El pre-ARNm,por ejemplo, sufre un proceso de maduración que tras cortes y empalmes sucesivos elimina ciertos segmentos del ADN llamados los intrones para producir el ARNm final. Durante este proceso de maduración se puede dar lugar a diferentes moléculas de ARN, en función de diversos reguladores. Así pues, un mismo gen o secuencia de ADN, puede dar lugar a diferentes moléculas de ARNm y por tanto, producir diferentes proteínas. Otro factor de regulación propio de las células eucariotas son los conocidos potenciadores (en inglés: "enhancers"), que incrementan mucho (100 veces) la actividad de transcripción de un gen, y no depende de la ubicación de éstos en el gen, ni la dirección de la lectura.
Etapas de la transcripción
Clásicamente se divide el proceso de la transcripción en 3 etapas principales (iniciación, elongación y terminación), pero realmente se pueden diferenciar 5 etapas :
Preiniciación
Al contrario de la replicación de ADN, durante el inicio de la transcripción no se requiere la presencia de un cebador para sintetizar la nueva cadena, de ARN en este caso. Antes del inicio de la transcripción se necesita toda una serie de factores de transcripción que ejercen los factores de iniciación. Estos se unen a secuencias específicas de ADN para reconocer el sitio donde la transcripción ha de comenzar y se sintetice el ARN cebador. Esta secuencia de ADN en la que se ensamblan los complejos de transcripción se llama promotor. Los promotores se localizan en los extremos 5'-terminales de los genes, antes del comienzo del gen, y a ellos se unen los factores de transcripción mediante fuerzas de Van der Waals y enlaces de hidrógeno. Los promotores tienen secuencias reguladoras definidas, muy conservadas en cada especie, donde las más conocidas son la caja TATA (situada sobre la región -10), con la secuencia consenso TATA(A/T)A(A/T); y la caja TTGACA (situada en el punto -35). La formación del complejo de transcripción se realiza sobre el promotor TATA, allí se forma el núcleo del complejo de iniciación. Sobre la caja TATA se fija una proteína de unión (TBP) junto con el factor de transcripción TFII D (TF proviene del inglés: transcription factor). Después, a ellos se unen otros factores de transcripción específicos: TFII B se une a TBP, TFII A (opcional), que estabiliza el complejo TFII B-TBP; luego se une el complejo TFII F y ARN polimerasa, y al final TFII E y TFII H. Todo ello forma un complejo que se llama complejo de preiniciación cerrado o PIC. Cuando la estructura se abre por mediación del factor de transcripción TFII H, da comienzo la iniciación y al complejo abierto (por su acción helicasa dependiente de ATP).
Iniciación
Primero, una Helicasa separa las hebras de ADN en estas denominadas cajas TATA, ya que entre adenina y timina se establecen dos enlaces de hidrógeno, mientras que entre citosina y guanina se forman tres. Posteriormente se unen los factores y las proteínas de transcripción (TBP, TF2D, TF2B) permitiendo, de esta manera, el acceso de la ARN polimerasa al molde de ADN de cadena simple, siendo esta la última en posicionarse. Aunque la búsqueda del promotor por la ARN polimerasa es muy rápida, la formación de la burbuja de transcripción o apertura del ADN y la síntesis del cebador es muy lenta. La burbuja de transcripción es una apertura de ADN desnaturalizado de 18 pares de bases, donde empieza a sintetizarse el ARN cebador a partir del nucleótido número 10 del ADN molde de la burbuja de transcripción. La burbuja de transcripción se llama complejo abierto. La ARN polimerasa es una enzima formada por 5 subunidades: 2 subunidades α, 1 subunidad β, 1 subunidad β' y 1 subunidad ω que tiene como función la unión de ribonucleótidos trifosfato. Cuando se forma el complejo abierto, la ARN polimerasa comienza a unir ribonucleótidos mediante enlaces fosfodiéster, y una vez que se forma el primer enlace fosfodiéster, acaba la etapa de iniciación y comienza así la siguiente etapa.
Disgregación del
promotor
Una vez sintetizado el primer enlace fosfodiéster, se debe deshacer el complejo del promotor para que quede limpio para volver a funcionar de nuevo. Durante esta fase hay una tendencia a desprenderse el transcrito inicial de ARN y producir transcritos truncados, dando lugar a una iniciación abortada, común tanto en procariontes como eucariontes. Una vez que la cadena transcrita alcanza una longitud de unos 23 nucleótidos, el complejo ya no se desliza y da lugar a la siguiente fase, la elongación.
La disgregación del promotor coincide con una fosforilación de la serina 5 del dominio carboxilo terminal de la ARN polimerasa, que es fosforilado por el TFII H (que es una proteína quinasa dependiente de ATP)
Elongación
La ARN polimerasa cataliza la elongación de cadena del ARN. Una cadena de ARN se une por apareamiento de bases a la cadena de ADN, y para que se formen correctamente los enlaces de hidrógeno que determina el siguiente nucleótido del molde de ADN, el centro activo de la ARN polimerasa reconoce a los ribonucleótidos trifosfato entrantes. Cuando el nucleótido entrante forma los enlaces de hidrógeno idóneos, entonces la ARN polimerasa cataliza la formación del enlace fosfodiéster que corresponde. A esto se le llama elongación, la segunda etapa de la transcripción del ARN.
Terminación
Al finalizar la síntesis de ARNm, esta molécula ya se ha separado completamente del ADN (que recupera su forma original) y también de la ARN polimerasa, terminando la transcripción. La terminación es otra etapa distinta de la transcripción, porque justo cuando el complejo de transcripción se ha ensamblado activamente debe desensamblarse una vez que la elongación se ha completado. La terminación está señalizada por la información contenida en sitios de la secuencia del ADN que se está transcribiendo, por lo que la ARN polimerasa se detiene al transcribir algunas secuencias especiales del ADN. Estas secuencias son ricas en guanina y citosina, situadas en el extremo de los genes, seguidas de secuencias ricas en timina, formando secuencias palindrómicas, que cuando se transcriben el ARN recién sintetizado adopta una estructura en horquilla que desestabiliza el complejo ARN-ADN, obligando a separarse de la ARN polimerasa, renaturalizándose la burbuja de transcripción. Algunas secuencias de ADN carecen de la secuencia de terminación, sino que poseen otra secuencia a la que se unen una serie de proteínas reguladoras específicas de la terminación de la transcripción como rho.
La traducción
es el
proceso por el cual la información genética contenida en el ADN y transcrita en
una ARN mensajero va a ser utilizada para sintetizar una
proteína de acuerdo a las reglas del código
genético. En este proceso intervienen los ARN ribosómico (ARNr)
y de transferencia (ARNt), y consta de las fases de:
-
activación: cada aminoácido (AA) se une al ARNt específico,
-
iniciación: la subunidad pequeña del ribosoma se enlaza con el extremo 5′ del
ARNm con la ayuda de factores de iniciación y de otras proteínas,
-
elongación: ocurre cuando el siguiente aminoacil-ARNt (el ARNt cargado) de la
secuencia se enlaza con el ribosoma,
-
terminación: sucede cuando se encuentra con un codón de terminación (sin
sentido), que son el UAA, UAG o UGA.
El código
genético es el conjunto de reglas usadas para traducir la
secuencia de nucleótidos del ARNm a una secuencia de proteína en el proceso de traducción.
Características del código genético:
• La correspondencia entre nucleótidos y aminoácidos se hace mediante codones. Un codón es un triplete de nucleótidos que codifica un aminoácido concreto.
• El código genético es degenerado: un mismo aminoácido es codificado por varios codones, salvo Triptófano y Metionina que están codificados por un único codón. Existen 64 codones diferentes para codificar 20 aminoácidos. Los codones que codifican un mismo aminoácido en muchos casos comparten los dos primeros nucleótidos.
• El codón AUG que codifica la metionina es el codón de inicio y hay tres codones que establecen la señal de terminación de la traducción (UAA, UAG, UGA).
• Es casi universal. Es el mismo para la mayoría de los organismos, con algunas excepciones en protozoarios, micoplasmas y las mitocondrias.
• La correspondencia entre nucleótidos y aminoácidos se hace mediante codones. Un codón es un triplete de nucleótidos que codifica un aminoácido concreto.
• El código genético es degenerado: un mismo aminoácido es codificado por varios codones, salvo Triptófano y Metionina que están codificados por un único codón. Existen 64 codones diferentes para codificar 20 aminoácidos. Los codones que codifican un mismo aminoácido en muchos casos comparten los dos primeros nucleótidos.
• El codón AUG que codifica la metionina es el codón de inicio y hay tres codones que establecen la señal de terminación de la traducción (UAA, UAG, UGA).
• Es casi universal. Es el mismo para la mayoría de los organismos, con algunas excepciones en protozoarios, micoplasmas y las mitocondrias.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}